The thin film hydration approach described here for the preparation of fluorescent liposomes enclosing drugs is a simple and cost-effective method. With the protocol used in this study, the liposomes are expected to be unilamellar23,24. The size, zeta potential, drug loading and entrapment efficiency of the liposomes produced are summarized in Table 1. The particle size of the liposomes (before and after drug loading) are similar (Table 1). The surface charge (zeta potential) of drug-loaded liposomes is slightly more neutral when compared with control liposomes, however, they are all negatively charged meaning this will not significantly change their biodistribution pattern.
Microinjection of Marina Blue-labeled liposomes into the hindbrain ventricle results in rapid uptake by resident macrophages that can be readily observed by confocal microscopy by 3 h post injection, when injected into the macrophage lineage-marking transgenic reporter line Tg(mpeg1:EGFP)33 (Figure 2C,E-G). This is in contrast to neutrophils (as marked within the neutrophil-specific Tg(lyz:EGFP)34 reporter line) that are rarely observed containing intracellular liposomes (Figure 2E-G). Within individual liposome-laden macrophages, the liposomes accumulate within phagolysosomal compartments (Figure 2H), which is necessary for liposome degradation and the subsequent release of their drug contents into the cytoplasm35. Selecting different microinjection sites for liposome delivery can impact which tissue-resident macrophages are targeted. As examples, delivery into the hindbrain ventricle efficiently targets hindbrain-resident macrophages (Figure 2D,E) while microinjection into the sinus venosus can deliver the liposomes to CHT-resident macrophages via the circulation (Figure 2I,J).
Consideration of the microinjection site for liposome delivery is important when using this technique to interrogate the macrophage response to an immunological challenge as it helps ensure the particular macrophages under investigation are receiving the drug. We have routinely used this protocol for the delivery of drug-loaded liposomes into the hindbrain ventricle to assess the impact of these drugs on the macrophage response to monosodium urate (MSU) crystals, similarly injected into the hindbrain compartment (Figure 3A). As the causative agent of acute gouty inflammation, MSU crystals activate tissue-resident macrophages to produce pro-inflammatory mediators including Interleukin-1β (IL-1β) through a process dependent upon mitochondrial reactive oxygen species (mROS)10,36,37,38. These activated macrophages then drive neutrophil infiltration, a hallmark of acute gouty inflammation. Microinjection of liposomes loaded with a mROS-inhibiting drug into the hindbrain ventricle can significantly suppress MSU crystal-driven mROS production within liposome-laden hindbrain-resident macrophages (Figure 3B-D). Using the protocol described here, we achieved an entrapment efficiency of 49.12 ± 0.17 %, which resulted in a formulation with a drug concentration of 103.05 ± 0.36 μM. Of note, injecting a 1 nL volume at this concentration resulted in no observable toxicity during our experiments, as evidenced by gross morphological changes or cardiac arrest. Further validation of the suppressive effects of this drug on macrophage activation state can be performed by investigating il1b expression (the zebrafish ortholog of IL-1β) by whole mount in situ hybridization (Figure 4A,B) and the temporal recruitment of neutrophils (Figure 4C,D).
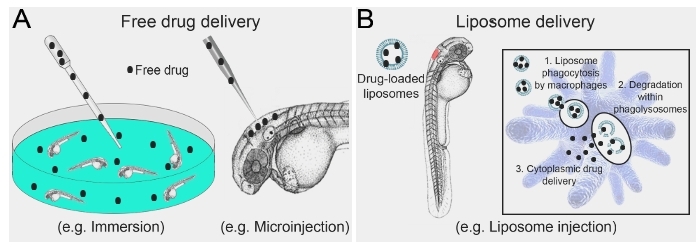
Figure 1: Schematic illustrating conventional free drug delivery versus liposome-mediated drug delivery to larval zebrafish. (A) Strategies routinely used for drug delivery to larval zebrafish are largely limited to immersion in, or microinjection of, free drug. (B) Microinjection of drug-loaded liposomes allows for direct targeting to macrophages, where liposome degradation within phagolysosomal compartments results in cytoplasmic drug delivery. Please click here to view a larger version of this figure.
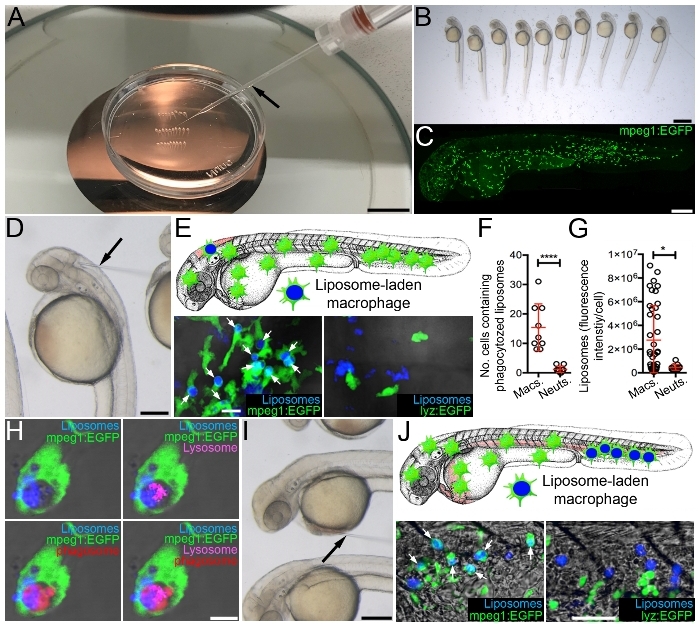
Figure 2: Targeting drugs to macrophages using fluorescent liposomes. (A) Microinjection set-up showing microinjection needle (black arrow) and larvae arrayed in a 35 mm tissue culture dish within 3% methylcellulose. (B) Magnified view of larvae arrayed as in A. (C) Live confocal image of 2 dpf Tg(mpeg1:EGFP) larvae, anterior to left. (D) Magnified view of arrayed larvae, as in A, demonstrating microinjection into the hindbrain ventricle (black arrow marks microinjection needle). (E) Schematic illustrating targeting of hindbrain-resident macrophages and live confocal images (dorsal views, anterior to left) of the hindbrain region of Tg(mpeg1:EGFP) and Tg(lyz:EGFP) larvae, 3 h following hindbrain microinjection of Marina Blue-labeled liposomes (white arrows mark liposome-laden macrophages). (F and G) Quantification of liposome uptake by macrophages and neutrophils (as detected in E), measured as the number of cells containing (F) and the fluorescence intensity/cell (G) of Marina Blue-labeled liposomes. (H) Live confocal image of liposome-laden macrophage within the hindbrain marked with a red fluorescent marker of phagosomes and a far red fluorescent marker of lysosomes within Tg(mpeg1:EGFP) larvae, 3 h following hindbrain microinjection of Marina Blue-labeled liposomes. (I) Magnified view of arrayed larvae, as in A, demonstrating microinjection into the sinus venosus (black arrow marks microinjection needle). (J) Schematic illustrating targeting of CHT-resident macrophages and live confocal images (lateral views, anterior to left) of the CHT region of Tg(mpeg1:EGFP) and Tg(lyz:EGFP) larvae, 3 h following microinjection of Marina Blue-labeled liposomes into the sinus venosus (white arrows mark liposome-laden macrophages). Error bars display mean ± SD. *p<0.05; ****p < 0.0001, Student's t-test. Scale bars = 100 mm (A), 100 μm (B), 250 μm (C, D, and I), 10 μm (E), 5 μm (H), 50 μm (J). This figure has been modified from previous publication24. Please click here to view a larger version of this figure.
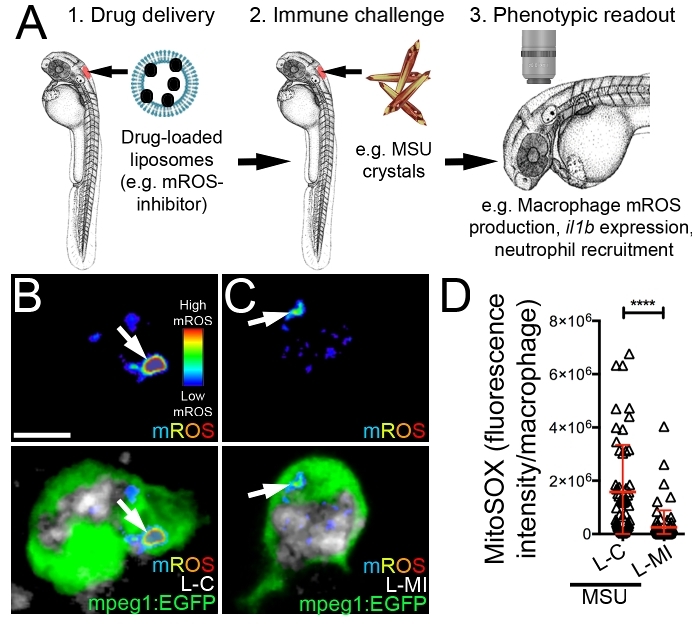
Figure 3: Microinjection of liposomes loaded with a mROS-inhibiting drug suppresses mROS production within activated macrophages. (A) Schematic illustrating the targeting of a mROS-inhibiting drug to macrophages and assessing it's impact on macrophage activation following MSU crystal stimulation. (B and C) Live confocal images of liposome-laden macrophages (control liposomes/L-C (B) or mROS-inhibiting liposomes/L-MI (C)) within the hindbrain of Tg(mpeg1:EGFP) larvae, also marked with a fluorescent mROS-specific probe (see Table of Materials, where the fluorescent signal is displayed as a heatmap with warm colors representing higher levels of mROS), 3 h following hindbrain microinjection of Marina Blue-labeled liposomes and MSU crystals. Marina Blue fluorescence is pseudo-colored in grayscale. (D) Quantification of fluorescence intensity of mROS-specific probe within macrophages, as detected in B and C. Error bars display mean ± SD. ****p < 0.0001, Student's t-test. Scale bar = 10 μm (B). This figure has been modified from previous publication24. Please click here to view a larger version of this figure.
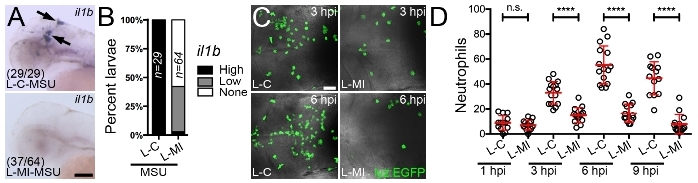
Figure 4: Microinjection of liposomes loaded with a mROS inhibitor suppresses il1b expression within activated macrophages and macrophage-driven neutrophil recruitment. (A) Expression of il1b (marked by black arrows), as detected by whole mount in situ hybridization, within the hindbrain region 3 h following hindbrain microinjection of control (L-C) or mROS-inhibiting liposomes (L-MI) and MSU crystals, anterior to left. Numbers represent proportion of larvae with displayed phenotypes. (B) Quantification of il1b expression, as detected in A, shown as percent larvae demonstrating high, low or no expression. (C) Confocal images of neutrophils within the hindbrain region of Tg(lyz:EGFP) larvae (dorsal views, anterior to left), as detected by immunofluorescence 3 (3 hpi) and 6 (6 hpi) h following hindbrain microinjection of L-C or L-MI and MSU crystals. (D) Temporal quantification of neutrophils, as detected in C, 1 (1 hpi), 3 (3 hpi), 6 (6 hpi) and 9 (9 hpi) h following hindbrain microinjection of L-C or L-MI and MSU crystals. Error bars display mean ± SD. ****p < 0.0001, n.s. not significant, one-way ANOVA, Dunnett's post hoc test. Scale bars = 100 μm (A), 50 μm (C). This figure has been modified from previous publication24. Please click here to view a larger version of this figure.
| Physicochemical property of liposomes (L) | Control liposomes | mROS-inhibiting liposomes |
| Size (μm) | 1.2 ± 0.07 | 1.1 ± 0.04 |
| Zeta potential (mV) | -25.9 ± 0.57 | -13.0 ± 0.11 |
| Entrapment efficiency (EE, %) | N/A | 49.12 ± 0.17 |
| Drug loading (DL, %) | N/A | 0.37 ± <0.01 |
Table 1: Physicochemical characteristics of PBS (control) or mROS-inhibiting liposomes (data are means ± standard deviation, n = 3). All liposomes were labeled with Marina Blue.
