Microglia is the CNS resident macrophage that gets activated when exposed to environmental challenges (trauma, toxic molecules, inflammation )4,5,6,34 (Figure 3A). In vitro studies on microglia are commonly used to evaluate cell-autonomous mechanisms related to those environmental challenges and characterize activation state after pharmacological or genetic manipulation. Here, an approach is presented to isolate primary microglia at the juvenile stage using magnetic coupled beads.
An easy readout for microglial activation in vitro is to quantify mRNA expression of several microglial reactivity markers associated with a pro-inflammatory phenotype (Cd86, Nos2, Ptgs2, Tnf) or an anti-inflammatory phenotype (Cd206, Igf1, Arg1, Lgasl3)5,6,31. These phenotypes were described depending on mRNA expression after pro-inflammatory (IL-1β, LPS) or anti-inflammatory (IL-4 or IL-10) stimulus. Some markers are classified as immuno-regulatory markers because they are upregulated through pro- and anti-inflammatory stimulation (Figure 3A). This classification was described previously by our team5,6,31. After 48 h, isolated microglial were stimulated with pro- and anti-inflammatory stimulus for 4 h or 24 h. mRNA were extracted, and RT-qPCR quantified gene expression. At 4 h and 24 h, pro-inflammatory and immuno-regulatory markers are induced by IL-1β, IL-1β + IFN-γ, and LPS5,6,31. At 4 h, IL-4 stimulation also induces the immuno-regulatory marker Il-1rn6. They are induced strongly at 4 h after IL-4 stimulation regarding anti-inflammatory markers. Interestingly, Cd206 is also significantly upregulated 24 h after IL-1β stimulation6 (Figure 3B).
To evaluate the phagocytic activity of microglia in vitro, fluorescent Cy3 PVC beads were used. They were pre-treated with fetal bovine serum (FBS) to facilitate their phagocytosis by microglia. Microglia were polarized toward pro-inflammatory phenotype by stimulation with IL-1β + IFN-γ or LPS for 3 h or 21 h. Three hours before the end of stimulation, Cy3-beads were incubated with microglial cells. After rinsing with 1x PBS, the fluorescence intensity in each well was quantified. Quantification relative to wells with no beads was expressed, and representative images were taken (Figure 4). After 6 h stimulation, microglia start to phagocyte Cy3-beads only under IL-1β + IFN-γ. After a 24 h stimulation, there is an increase of Cy3 fluorescence for both kinds of stimulation. That increase of Cy3 fluorescence highlights increased phagocytic activity (Figure 4C).
Flow cytometry (FACS) and RT-qPCR were performed to evaluate the microglial culture's purity. Different brain cellular populations can be distinguished by flow cytometry: CX3CR135 for microglia, O4 for oligodendrocytes36, NeuN for neurons37, and ACSA-2 for astrocytes38. After dissociation, all brain cells are present; however, after cell sorting using CD11b antibody, only microglia and small amounts of O4 cells are present (Figure 5A). 48 h after primary cell culture, only microglia markers are found as evaluated by RT-qPCR (Figure 5B).
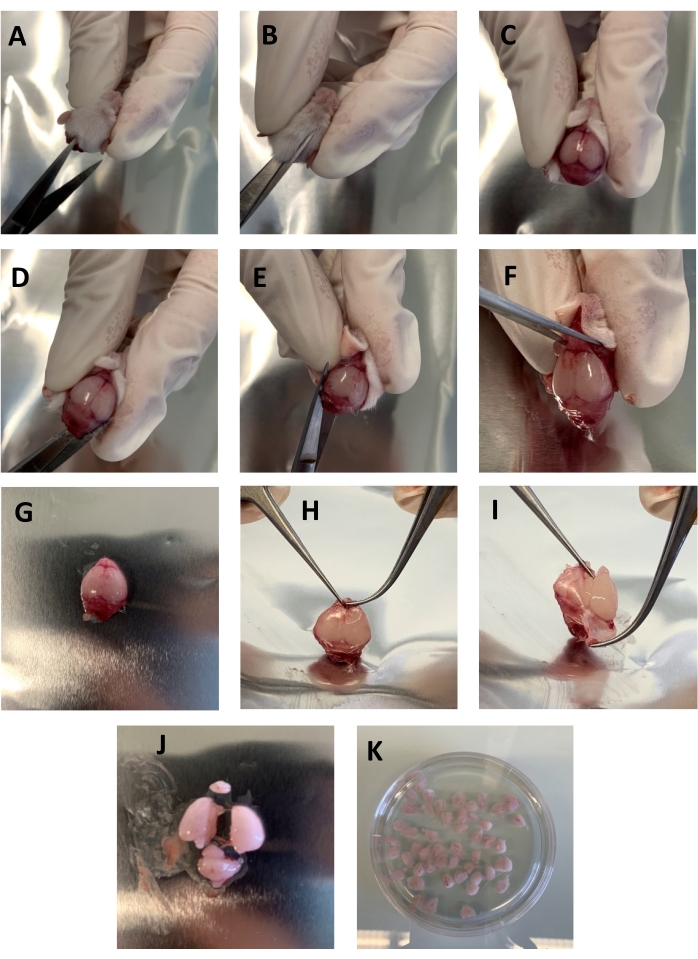
Figure 1: Step-by-step representation showing the removal of the brain from the skull. (A–C) Small scissors were used to cut the skin from the neck to the nose following the sagittal suture. (D–E) The small scissor's tips were inserted within the Foramen magnum parallel to the skull, and from each side to the eyes were carefully cut. (F) The skull and brain were detached from the head by cutting between the eyes with small scissors. (G–I) The skull was grabbed close to the olfactive bulbs with two forceps and torn carefully not to damage the underlying brain. (J) The cerebellum and the olfactive bulb were removed with a razor blade, and the brain was cut into two pieces. (K) The brain pieces were placed in a Petri dish containing 30-40 mL of HBSS-/-. Please click here to view a larger version of this figure.
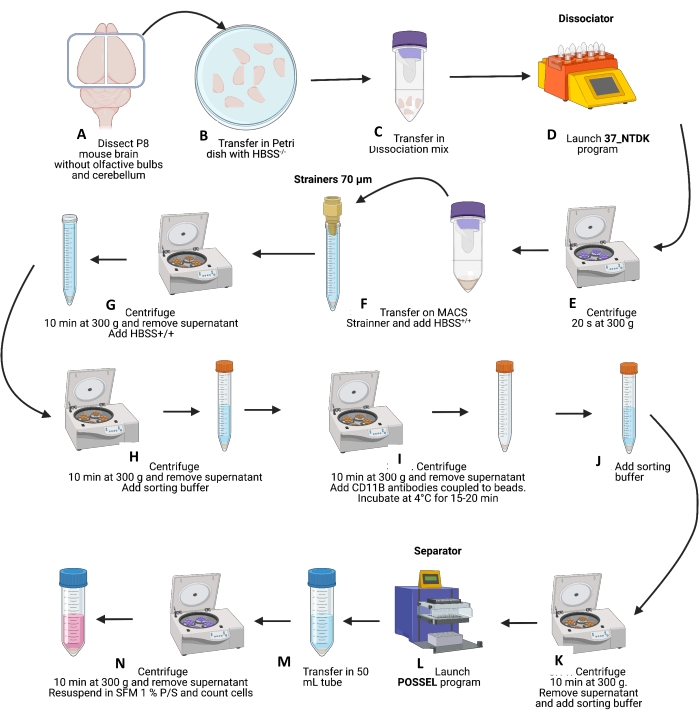
Figure 2: Schematic representation of brain dissociations and microglial cells isolation. (A–C) After P8 mouse brain dissection and removing olfactive bulbs and cerebellum, brains were first transferred to a Petri dish with HBSS+/+, and then to dissociation tubes containing the dissociation mixture. The C-tubes were placed on the dissociator (with the heating mode), and the NTDK program was started (D). (E) At the end of the program, tubes were centrifuged at 300 x g for 20 s at 4 °C; dissociation was then completed by pipetting three times up and down with a 1,000 µL pipette. (F) Cells were then transferred to 15 mL tubes + strainers of 70 µm and rinsed with 10 mL of HBSS+/+. (G) Samples were then centrifuged at 300 x g for 10 min at 4 °C, the supernatant was removed, and the pellet was resuspended with 10 mL of HBSS+/+. (H) Tubes were again centrifuged at 300 x g for 10 min at 4 °C; the supernatant was removed, and then the pellet was resuspended in 6 mL of sorting buffer. (I) Tubes were centrifuged at 300 x g for 10 min at 4 °C, the supernatant was removed, and the CD11b-microbead (200 µL) solution was added. Tubes were incubated for 15-20 min at 4 °C, and then resuspended in 6 mL of sorting buffer (J) and centrifuged at 300 x g for 10 min at 4 °C. (K) The supernatant was removed, and the pellet was carefully resuspended in 8 mL of sorting buffer. (L) Then, launch the POSSEL program on the separator to prepare columns. Cells were transferred 1 mL by 1 mL on the column, and CD11b cells were eluted on a sterile elution plate with 1 mL of sorting buffer. (M) CD11b cells were pooled in a new 50 mL tube and centrifuged at 300 x g for 10 min at 4 °C, and the supernatant was removed. (N) The pellet was carefully resuspended in 10 mL of cold microglia medium in the last step. The cells were counted and diluted in the microglial medium to a final concentration of 650,000-700,000 cells/mL dispensed in cell culture plates. Please click here to view a larger version of this figure.
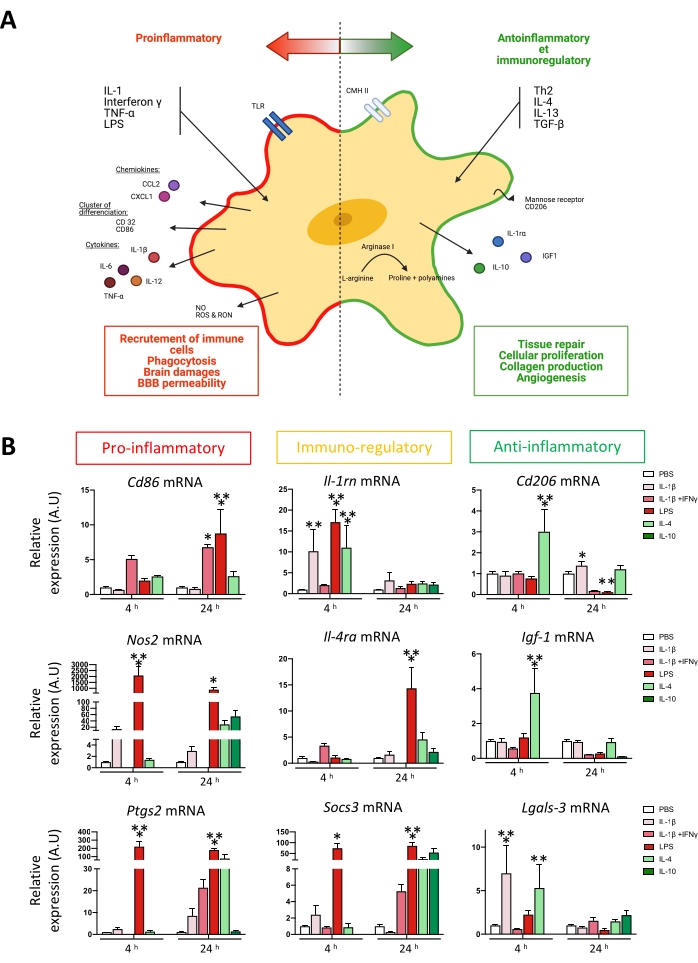
Figure 3: Microglial activation following exposition to environmental challenges. (A) Simplified schematic representation of microglial activation spectrum. (B) Relative quantification of microglial activation markers after 4 h or 24 h stimulation. Two-way ANOVA followed by Dunnett's multiple comparisons test39 (n = 5-15); error bars represent SEM; *p < 0.05, **p < 0.01, ***p < 0.001. Please click here to view a larger version of this figure.
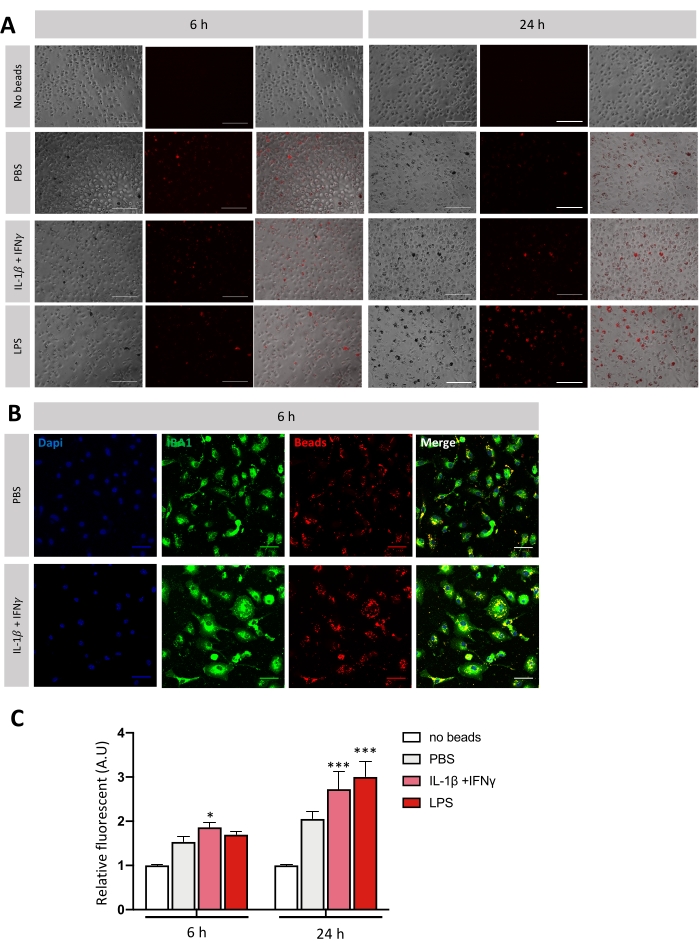
Figure 4: Evaluation of the phagocytic activity of microglia in vitro. (A) Representative images of microglia Cy3-beads (in red) phagocytosis after stimulation of 6 h or 24 h. The images were acquired using 20x objective of the fluorescence microscope. Scale bar = 100 μm. (B) Relative quantification of fluorescence emits per well. Statistics was performed using two-way ANOVA followed by Dunnett's multiple comparisons test (n = 4-7); error bars represent SEM; *p < 0.05, ** p < 0.01, ***p < 0.001. (C) Representative picture of microglia (IBA-1 + cells in green) Cy3-beads (in red) phagocytosis after 6 h stimulation. The images are acquired using a 40x objective of the confocal microscope. Scale bar = 300 μm. Please click here to view a larger version of this figure.
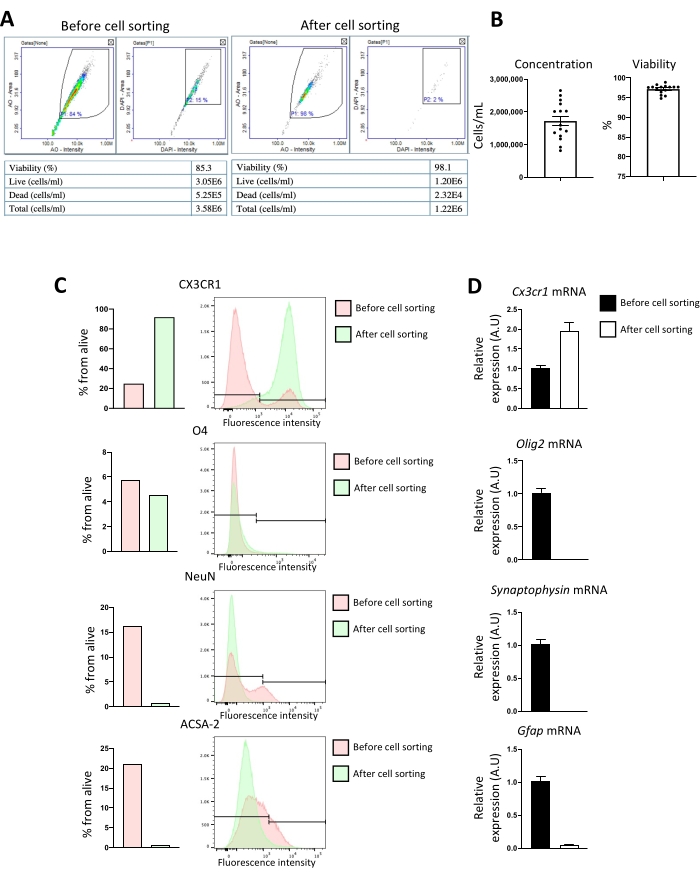
Figure 5: Evaluation of CD11b magnetic isolation purity by flow cytometry before and after cell sorting. (A) Reports examples from the cell counter before and after cell sorting, highlighting cell concentration and viability. (B) The x-axis represent cell concentration (cell/mL) and viability (%) after CD11b cell sorting. Column bar graph; error bars represent SEM (n = 16). Phenotypic and gene expression analysis of cell population markers before and after CD11b+ cell sorting. After dissociation, CD11b+ cells from P8 mice brains were magnetically sorted. Expression of microglia (CX3CR1), oligodendrocytes (O4 or Olig2 mRNA), neurons (NeuN or Synaptophysin mRNA)), and astrocytes (ACSA-2 or Gfap mRNA) markers was analyzed before and after sorting. (C) FACS analysis of CX3CR1, O4, NeuN, and ACSA-2 expression. The x-axis represents the percentage of alive cells and cell numbers. (D) Relative quantification of CD11b+ cells expression of Cx3cr1, Olig2, Synaptophysin, and Gfap mRNA (x-axis represent relative target mRNA expression to Itgam mRNA). Column bar graph; error bars represent SEM (n = 7). Please click here to view a larger version of this figure.
| Solution | For one C-Tube (µL) | For four C-tubes (µL) |
| Buffer X | 2850 | 11400 |
| Enzyme P (Papain) | 75 | 300 |
| Enzyme A (DNAse) | 15 | 60 |
| Buffer Y | 30 | 120 |
| Total | 2970 | 11880 |
Table 1: Preparation of the dissociation mixture.
| Solution | For one tube (µL) | For four tubes (µL) |
| CD11b Microbeads | 20 | 80 |
| Sorting Buffer | 180 | 720 |
| Total | 200 | 800 |
Table 2: Preparation of the CD11b microbead solution.
| Stimulation | Concentration (ng/mL) |
| IL-1β | 50 |
| IFN-γ | 20 |
| LPS | 10 |
| IL-4 | 30 |
| IL-10 | 20 |
Table 3: The stimulation reagents.
| Genes | Protein | Forward | Reverse |
| Arg1 | Arginase 1 | GTG AAG AAC CCA CGG TCT GT | GCC AGA GAT GCT TCC AAC TG |
| Cd206 | Cluster of differentiation 206 | CTT CGG GCC TTT GGA ATA AT | TAG AAG AGC CCT TGG GTT GA |
| Cd32 | Cluster of differentiation 32 | CTG GAA GAA GCT GCC AAA AC | CCA ATG CCA AGG GAG ACT AA |
| Cd86 | Cluster of differentiation 86 | GAG CGG GAT AGT AAC GCT GA | GGC TCT CAC TGC CTT CAC TC |
| Gfap | Glial fibrillary acidic protein | AAGCCAAGCACGAAGCTAAC | CTCCTGGTAACTGGCCGACT |
| Igf-1 | Insulin like growth factor 1 | TGG ATG CTC TTC AGT TCG TG | GCA ACA CTC ATC CAC AAT GC |
| Il1-rn | Interleukine 1 receptor antagonist | TTG TGC CAA GTC TGG AGA TG | TTC TCA GAG CGG ATG AAG GT |
| Il4-ra | Interleukine 4 receptor antagonist | GGA TAA GCA GAC CCG AAG C | ACT CTG GAG AGA CTT GGT TGG |
| Itgam | Integrin alpha M | CTGGTGCTCTTGGCTCTCAT | GGCAGCTTCATTCATCATGT |
| Lgals3 | Lectin Glactoside-Binding soluble 3 | GAT CAC AAT CAT GGG CAC AG | ATT GAA GCG GGG GTT AAA GT |
| Nos2 | Nitric oxide synthase 2 | CCC TTC AAT GGT TGG TAC ATG G | ACA TTG ATC TCC GTG ACA GCC |
| Olig2 | Oligodendrocyte transcription factor 2 | CAGCGAGCACCTCAAATCTA | GATGGGCGACTAGACACCAG |
| Ptgs2 | Prostaglandin endoperoxide synthase 2 | TCA TTC ACC AGA CAG ATT GCT | AAG CGT TTG CGG TAC TCA TT |
| Rpl13 | Ribosomal protein L13a | ACA GCC ACT CTG GAG GAG AA | GAG TCC GTT GGT CTT GAG GA |
| Socs3 | Suppressor of cytokine 3 | CGT TGA CAG TCT TCC GAC AA | TAT TCT GGG GGC GAG AAG AT |
| Sphk1 | Sphingosine kinase 1 | TCC AGA AAC CCC TGT GTA GC | CAG CAG TGT GCA GTT GAT GA |
| Syp | Synaptophysin | ATCTCAGTGTCCCGATCCCA | GCTGTCTTCCTGGTGGGTAC |
| Tnf-α | Tumor necrosis factor α | GCC TCT TCT CAT TCC TGC TT | AGG GTC TGG GCC ATA GAA CT |
Table 4: RT-qPCR primer sequences.
| Stimulation | For 1-well | For n-wells |
| Cy3 Beads (1 cell : 50 beads) | 8, 100, 000 | X = (8, 100, 000) x n |
| 1x PBS | Y =(100+X)/n | 50 |
| FBS | 50 |
Table 5: Preparation of the beads mixture for phagocytic assay.
