Representative data are presented here to demonstrate successful intraductal injection, successful viral infection, and the impact of the delivered genes on mammary tumorigenesis. The amount of virus injected must be tailored to the purpose of each experiment. To illustrate how extensively the mammary duct tree can be infected, a large amount of virus carrying genes that can be imaged, such as GFP, needs to be used. On the other hand, to mimic the natural spontaneous tumorigenesis, a small amount of virus carrying an oncogene must be used so that only a few cells are infected and will evolve to precancerous lesions, and eventually invasive cancer, in a field of the otherwise completely normal mammary gland.
The success of intraductal injection can be immediately confirmed by exposing the mammary gland and observing a blue ductal tree (Figure 1). Two to five days after injection of the Lenti-EGFP (FUCGW) virus (~106 IUs)12, mammary epithelial cell infection can be assessed by whole mount preparation followed by observing under a fluorescent stereomicroscope (Figure 2). Alternatively, for flow cytometry analysis of the fluorescent proteins or cell surface markers produced by the virus6,7,19, the infected glands and non-infected glands (as negative controls) may be collected and processed into a single-cell suspension so that the rate of viral infection can be estimated (Figure 3). Another method to test/quantify virus infection is to fix the infected and non-infected control glands using 4% paraformaldehyde (or formalin), process them into paraffin-embedded blocks, and stain the resulting sections for the virally produced gene products and epitope tags such as HA and FLAG20,21.
Whether the virally infected cells will expand and form precancerous lesions, and then tumors, depends on the potency of the oncogene(s) delivered and the amount of virus injected. For a potent oncogene such as PyMT and activated RAS, early lesions form in a few days, and tumors appear in a few weeks10 (Bu et al., unpublished observations). Activated ERBB2 leads to precancerous lesions in a few weeks and tumors in a few months10,22,23 (Figure 4), while activated PIK3CA leads to tumors with a median latency of approximately 5 months24. On the other hand, Wnt1 causes tumors rather slowly: only about 20% of the infected mice developed tumors after 20 months21.
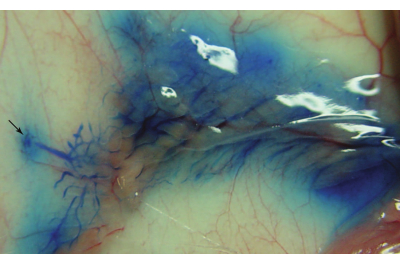
Figure 1: A successfully injected mammary ductal tree. The image was captured immediately after the intraductal injection of the number 4 mammary gland of a 9-week-old FVB/N female mouse. The arrow indicates the location of the number 4 nipple. Please click here to view a larger version of this figure.
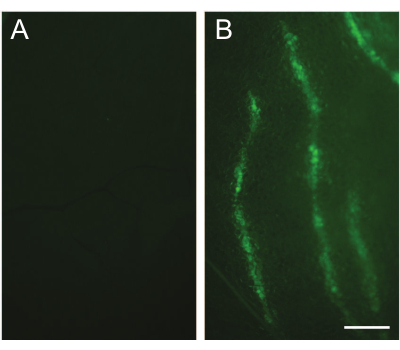
Figure 2: Detection of infected cells by fluorescent stereomicroscopy. (A) An un-injected contralateral gland is used as a control. (B) An image captured under a fluorescent stereomicroscope shows the infected mammary duct tree. The number 3 mammary gland of a 10-week-old FVB/N mouse was intraductally injected with the Lenti-EGFP (FUCGW) virus (~106 IUs). The injected mammary gland was collected 5 days after the injection. Scale bar: 1 mm. Please click here to view a larger version of this figure.
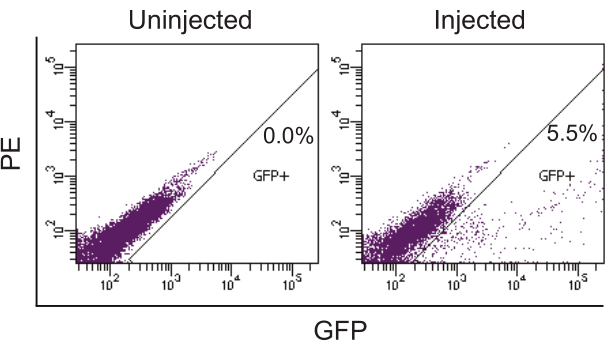
Figure 3: Quantifying infected cells by flow cytometry analysis for EGFP produced by lentivirus. The phycoerythrin (PE) channel was used to reveal the auto-fluorescent signal. The contralateral uninjected mammary glands were used as the negative control. The injected mammary glands from a 12-week-old FVB/N female mouse were collected 2.5 days after intraductal injection of Lenti-EGFP (~106 IUs/gland) and processed into a single-cell suspension. The single-cell suspension was then analyzed by flow cytometry to detect the GFP-positive cells. Please click here to view a larger version of this figure.
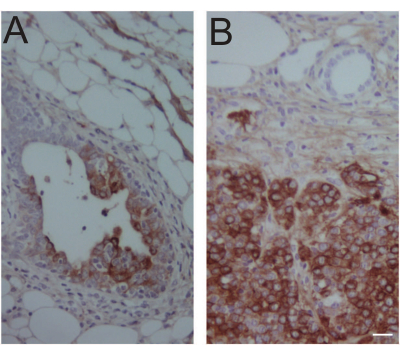
Figure 4: Immunohistochemical confirmation of virus-expressed oncogene product in an early lesion and a tumor. (A) An early lesion-containing #4 mammary gland was collected 14 days after intraductal injection of 106 IUs of RCAS-Neu (HA) into MMTV-tva mice. Immunohistochemical staining was used to detect the HA tag. (B) A tumor was collected 1 year after intraductal injection of 104 IUs of RCAS-Neu (HA) into a #4 gland of MMTV-tva mice. Mouse age at injection: 12 weeks. Scale bar: 20 µm. Please click here to view a larger version of this figure.
