The animals were trained and tested with four female rats in one colony cage and four male rats in a separate colony cage. All rats learned to pass through the ORTs in four days or less. The four female rats reached >85% successful bouts at the 120 g force requirement in approximately 6 weeks of training and the male rats reached the same criterion in 10 weeks (compared to roughly 3 weeks with standard training with deprived rats)7. This training duration was greatly lengthened due to several hardware and software glitches that required continuous troubleshooting, between weeks 2 to 6. Once these glitches were addressed, training proceeded smoothly, and it is expected that subsequent training timelines will be comparable to the current literature7. The male rats were also trained longer to provide more opportunities for one male rat to begin pulling; however, he never did and was excluded from further analysis and surgery after week 7. Once rat performance steadied in baseline, 5 days of pre-stroke baseline data was obtained for each cage. The data was limited to days during which the ORT remained connected to the cage for the entire day (some days required husbandry-related temporary disconnections). For the female cage, baseline days were 7, 8, 9, 10, and 12 days before stroke. For the males, baseline days were 8, 9, 10, 11, and 13 days before stroke.
During their stroke induction surgeries, the animals in this validation were simultaneously implanted with electrodes connected to wireless receiving chips in either their Basal Forebrain (coordinates -5.8 mm anterior/posterior, 0.7 mm left medial/lateral, 8.3 mm dorsal/ventral) or their Ventral Tegmental Area (coordinates -2.3 mm anterior/posterior, 3.3 mm left medial/lateral, 7.0 mm dorsal/ventral). These implants were for use in a subsequent recovery experiment and are not relevant to the colony cage-ORT validation reported here. The implants were designed so that the skin could be closed over them, with the receiving chips located subcutaneously under the left arm.
One female animal died during stroke induction. Another female began to decline several days after recovery, having never pulled the handle after stroke. After her euthanasia, it was found that she had probably experienced a brain hemorrhage sometime after her stroke. These two animals were eliminated from the data set entirely, including the pre-stroke assessment.
After stroke, animals did not immediately reinitiate regular lever pulling, though they continued to enter the chamber via the ORT and had to be encouraged via short sessions of manual shaping (i.e., reduced lever distance and delivered rewards contingent upon approaching or attempting to pull the lever). Female rats did not pull during days 4-7 after stroke and so were given supplementary lever baiting (i.e., a dab of peanut butter on the lever) and manual shaping on days 8-11. They began pulling autonomously on day 11. Males were allowed to recover until day 6 based on previous experience with the females. They did not pull during day 6 after the stroke. They were given supplementary baiting on day 7. They began autonomously pulling on the 8th day after the stroke. Once the animals again contacted reinforcement for pull attempts, supplementary baiting or shaping was ceased and post-stroke data was gathered. Males did not pull enough on day 8 for complete analysis of the more complex dependent variables (circadian measures and post-bout pausing), so they were allowed to continue to pull on the 9th, 10th, and 11th day under the same criterion. The 8th, 10th, and 11th day were complete days. The first day of pulling after stroke was used for all analyses except the circadian analysis and the analysis of pauses between bouts; for this analysis the one day for females and the three complete days for the males were used. For post-stroke analysis, the two female rats provided 55 and 844 pulls in one day, and the three male rats provided 536, 153, and 190 pulls in three days.
Data were organized according to pulls and bouts. To avoid registering tremors arising from the equipment itself, pulls were measured using a 5 g threshold with a +/- 1 g hysteresis. A pull was registered when the animal exerted pressure above 6 g and stopped when the handle registered a force below 4 g. Animals tended to pull in bouts of several rapid pulls. Once any single pull reached 120 g, reinforcement was delivered. A bout was considered a cluster of pulls whose peaks were all separated by less than 1 s. This threshold was selected based on previous data, which indicated that an inter-peak interval cluster under 1 s naturally developed, and other inter-peak intervals were reliably much longer. Rats would generally pull many times in a row before visiting the feeder, even when earlier pulls in the bout activated the feeder.
A total of 7 dependent variables were analyzed. A paired t-test was performed between baseline averages and post-stroke measures, which are reported in Figure 1, Figure 2, and Figure 3. These figures also display data by individual animals to provide an impression of the variation across days and between individuals for each measure that might be expected.
Figure 1 shows pre- and post-stroke performance along several performance measures typical of skilled reach assessments7,8,10. All post-stroke data was aggregated into a single data point even if it took several days to collect sufficient trials. The protocol and automated self-initiated system successfully assessed success rate per bout, mean force per pull, and pulls per bout, which all showed sensitivity to the stroke with varying degrees of statistical significance.
Figure 2 depicts two novel variables that arise from the colony cage-ORT setup: session initiations and cumulative session duration. Surprisingly, stroke did not affect session initiations. Females reliably initiated sessions more than males both before and after stroke, however neither changed their rate after stroke. Conversely, duration spent in the chamber increased for most rats, perhaps due to the decreased success rate of bouts (the result of which is a decreased rate of rewards)
Session initiation (which represents a choice between enrichment and social rewards available in the colony cage and food reinforcement) and duration of time in the chamber (in the case of a conditioned place preference with the value of reward) could also be taken as indices of motivation24,25,26,27. Additional motivation-based measures were included, such as "effort" as quantified by pulls per session minute28 and pauses between bouts29, which can be seen in Figure 3. These variables were impacted by stroke. As anticipated, the number of pulls per session minute decreased, and the duration of pauses between bouts increased. However, changes in the latter measure were complex. The distribution of bout pauses seemed to become more chaotic, including more long pauses, a few very long pauses, and more short pauses as well. This may indicate a breakdown in the original skilled motor unit; if so, it might be an easily measurable index of the same.
Despite the small group size, an investigation was conducted to determine if any of the measured variables exhibited correlations with the success rate, potentially implying their functional significance. Shapiro Wilk tests were performed to assess for equal distributions of the data for the variables success rate, mean peak pull, bout per minute, cumulative session duration, pauses between bouts, and pulls per bout. The Shapiro Wilk test indicated that the distribution of some variables departed significantly from normality. Therefore, Spearman's rank-order correlations were performed to determine the relationship between pre- or post-stroke success rate and the following variables: mean peak, pull bout per minute, cumulative session duration, pauses between bouts, and pulls per bout. No variables before stroke other than mean pull force were meaningfully correlated with success rate (see Table 1). After stroke, most variables also showed no meaningful correlation with success rate, except mean pull force (Table 1).
Finally, the ORT permits an analysis not only of skilled motor behavior, but also of the circadian patterning. Figure 4 shows the proportion of each hour for each cage in which the ORT was occupied, presented as an average over baseline and post-stroke days. The blue line in the figure indicates an averaged count of the number of entries per hour made throughout the day. Before stroke, animals would engage with the skilled reach task at high levels in the morning, diminishing their durations throughout the day. A few hours before lights-on, engagement either spiked again (females) or increased very slightly (males) before discontinuing shortly after the lights came on. This bimodal circadian distribution completely changed after stroke. Animals engaged less in the mornings, and their time in the chamber peaked later in the day. Such patterning may be reflective of the general sleep and circadian disturbances often observed after stroke30,31,32,33.
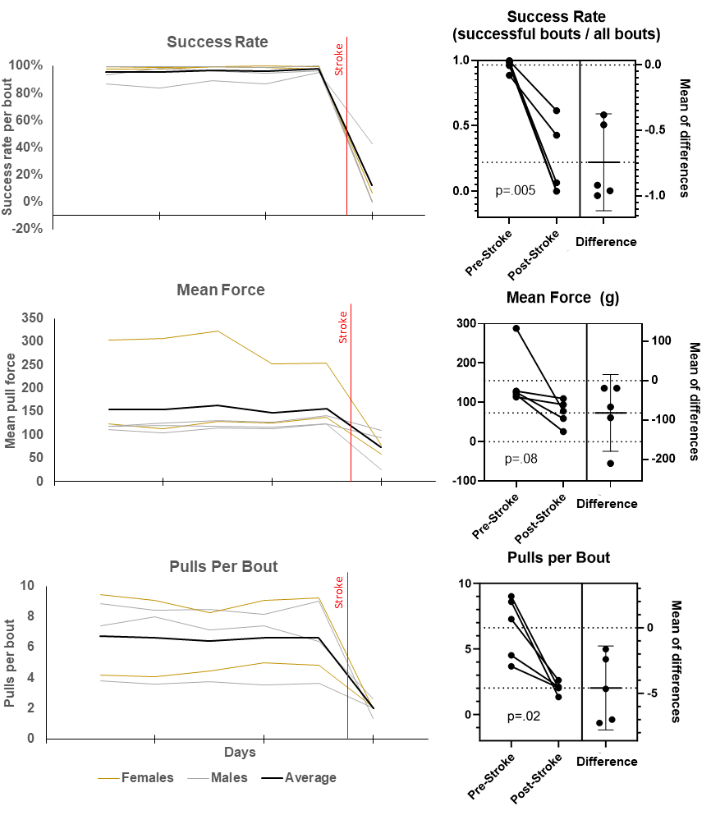
Figure 1: Measuring the typical performance changes in skilled reach after stroke using the ORT procedure. The performance of skilled reach before and after stroke was measured. Daily averages by animal for success rate per bout, mean force per pull, and number of pulls per bout are displayed over 5 days of baseline and one day post-stroke (left) and between an average of baseline and one day post-stroke (post-stroke) with reported paired t-tests. Please click here to view a larger version of this figure.
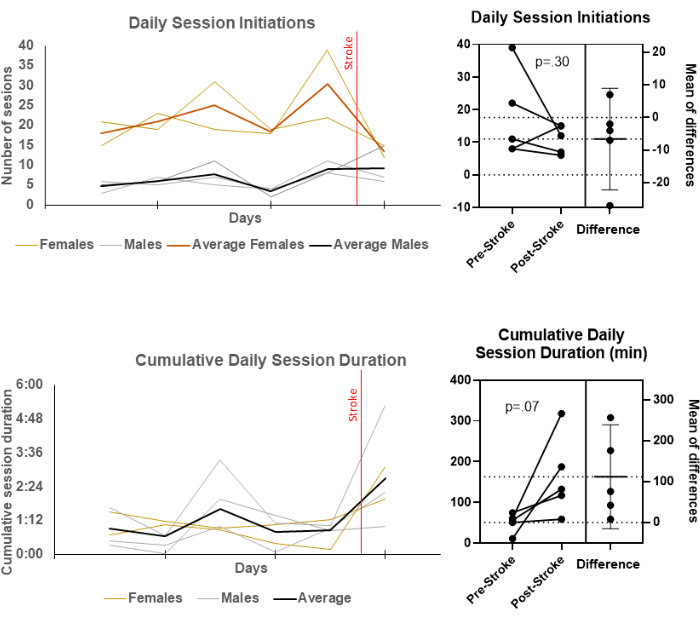
Figure 2: Different self-initiated session parameters show different tendencies to change after stroke. The measures of self-initiation for skilled reach behavior sessions before and after stroke was made possible by the ORT procedure. Daily averages by animal for session initiations and cumulative daily session durations are displayed over 5 days of baseline and one day post-stroke (left) and between an average of baseline and one day post-stroke (post-stroke) with reported paired t-tests. Please click here to view a larger version of this figure.
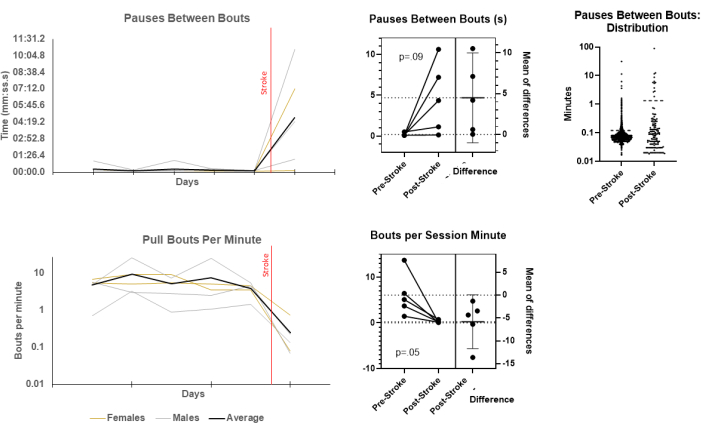
Figure 3: Motivation-related variables change after stroke. The performance measures of skilled reach behavior sessions before and after stroke related to motivation was determined. Daily averages by animal for pause duration between bouts and daily rate of bouts per session minute are displayed over 5 days of baseline and one day post-stroke (left) and between an average of baseline and one day post-stroke (post-stroke) with reported paired t-tests. Pauses between bouts changed in terms of these daily averages, but even more strikingly, the distribution of individual pause lengths after stroke also changed on both sides of the average. Individual pause lengths are pooled for all animals and displayed as distributions on a log axis (far right). Please click here to view a larger version of this figure.
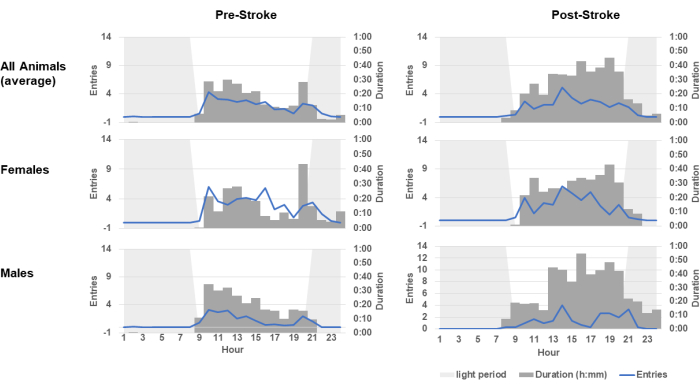
Figure 4: Circadian patterning of self-initiated sessions changes after stroke. The measures of circadian patterning of self-initiated skilled reach behavior sessions before (left) and after (right) stroke for all animals, females, and males were determined. These data include all entries and all times of chamber occupancy by cage, averaged across pre-stroke and post-stroke days. The two cages are then averaged again to show total distributions (top row). Pre-stroke patterns included high engagement in the morning, which diminished throughout the waking period with a new peak just before sleep phase. Post-stroke patterns show session durations increasing through the day and peaking before the sleep phase. Rats were on reverse light cycles in their housing room. The lights-on period is shown shaded in grey to indicate the rat's normal inactive period. Please click here to view a larger version of this figure.
| Correlations | ||||||
| Variables | Comparison Variable | Spearman | ||||
| n | Before Stroke rs | p | After Stroke rs | p | ||
| Mean Pull Force | Success Rate | 5 | 1 | <0.001 | -0.975 | 0.005 |
| Pull Bout per Minute | Success Rate | 5 | 0.3 | 0.624 | -0.154 | 0.805 |
| Cumulative Session Duration | Success Rate | 5 | -0.1 | 0.873 | 0.564 | 0.322 |
| Pauses Between Bouts | Success Rate | 5 | -0.6 | 0.285 | 0.205 | 0.741 |
| Pulls per Bout | Success Rate | 5 | 0.1 | 0.873 | -0.821 | 0.089 |
Table 1: Spearman correlation coefficients between variables. Spearman's rank-order correlations were performed to determine the relationship between pre- or post-stroke success rate and the following variables were determined: mean peak, pull bout per minute, cumulative session duration, pauses between bouts, and pulls per bout. Prior to the correlation, Shapiro Wilk tests were performed to assess for equal distributions of all test variables success rate, and indicated some variables departed significantly from normality. No variables before stroke other than mean pull force were meaningfully correlated with success rate. This table depicts the results of Spearman correlation coefficients (ρ) assessed if there was a relationship between success rate and five test variables.
Supplementary File 1: Steps to building the ORT. Instructions for printing and constructing a "One Rat Turnstile." Included in the instructions are a list of all needed materials as well as step-by-step instructions (with images). The file also includes directions for attaching a microswitch to register entries and exits, as well as the wiring and programming for attaching an RFID-reader. Please click here to download this File.
Supplementary Coding File 1: This includes all the components needed for 3D-printing the "One Rat Turnstile." This file can either be used directly or accessed using the instructions in Supplementary File 1. All components in this file must be scaled using the "ruler" piece that is included (see Supplementary File 1 for further details). Please click here to download this File.
