The tissue best suited for protoplast isolation is the second and third leaves of 10-11 day old seedlings (Figure 1). In this work, we obtained approximately 10 million protoplasts from 16 leaf sections, each of which was 10 cm long. The number of protoplasts isolated is dependent on the mass of the leaf material and the width of the leaf slices that are digested in the enzyme solution.
Under a brightfield light microscope, undamaged protoplasts appear roughly spherical, with plastids speckling the surface (Figure 2A). Before proceeding to transfection, the viability can be checked by staining a small aliquot of protoplasts with fluorescein diacetate (FDA). FDA-stained protoplasts that fluoresce under UV light are considered viable (Figure 2B,C). Not all protoplasts that appear undamaged are still viable, and some viable protoplasts may be in the process of dying by evacuolation (Figure 2C). In these cells, the vacuole is swelling and will eventually be ejected from the cell membrane.
Figure 3 shows protoplasts transformed with pJT01, a 4.3 kb maize expression plasmid derived from CD3-911 (ABRC)15. Transfection results in the expression of sGFP tagged with a C-terminal nuclear localization signal from c-Myc (Figure 3A). The proportion of sGFP-expressing protoplasts was measured over multiple experiments using a hemocytometer. The median transfection efficiency achieved by this method was 40% after overnight incubation (n = 9, Figure 4, Table 1), which is comparable to previously published methods15. Depending on the experimental application, if the plasmid or plasmid library lacks a fluorescent reporter, it is recommended to include a positive control to confirm transfection.

Figure 1: Representative etiolated maize seedling grown in darkness for 10 days after germination. The arrows indicate the second and third leaves, which are best suited for protoplasting. Please click here to view a larger version of this figure.
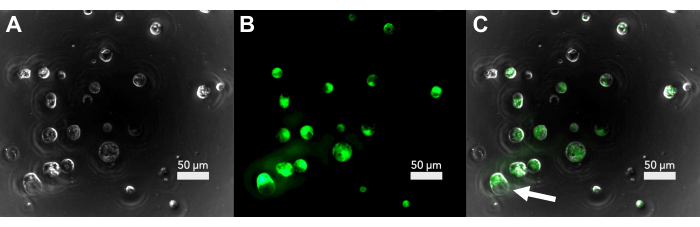
Figure 2: Fluorescence microscopy of the protoplast viability after isolation. (A) Brightfield image of protoplasts immediately after isolation. (B) The fluorescence signal of the FDA-stained protoplasts. (C) Overlap of brightfield and fluorescence showing the viable protoplasts. The arrow shows a protoplast undergoing evacuolation. Please click here to view a larger version of this figure.
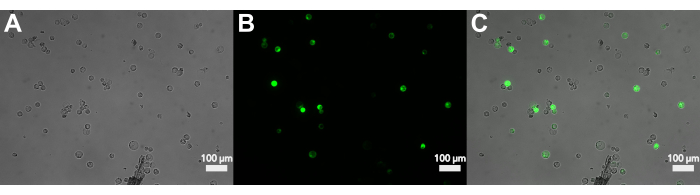
Figure 3: Fluorescence microscopy of protoplasts transfected with sGFP. (A) Brightfield image of protoplasts after transfection. (B) The fluorescence signal of the transfected protoplasts in the GFP channel. (C) Overlap of brightfield and fluorescence showing the transformed protoplasts. Please click here to view a larger version of this figure.
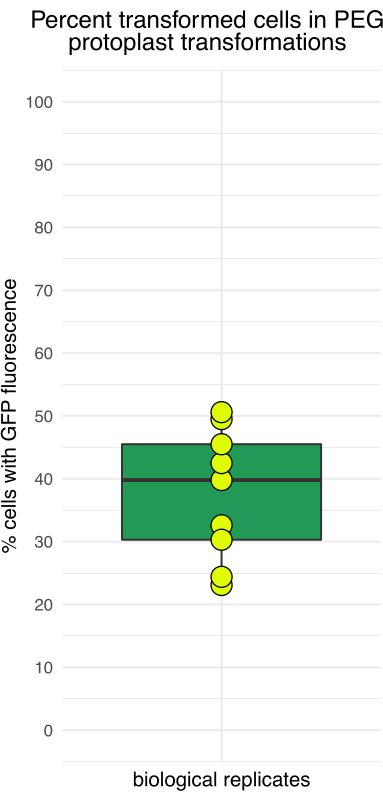
Figure 4: Percentage of protoplasts showing GFP fluorescence after 16 h of incubation. The transfection efficiency varied among independent trials, with the lowest efficiency around 20% and the highest close to 50%. Please click here to view a larger version of this figure.
| Trial | GFP cells counted | Total cells counted | GFP percentage |
| 1 | 104 | 261 | 39.8 |
| 2 | 148 | 298 | 49.5 |
| 3 | 84 | 258 | 32.6 |
| 4 | 137 | 271 | 50.6 |
| 5 | 127 | 299 | 42.5 |
| 6 | 107 | 235 | 45.5 |
| 7 | 83 | 360 | 23.1 |
| 8 | 134 | 549 | 24.4 |
| 9 | 61 | 201 | 30.3 |
| Mean | 109 | 304 | 36 |
| Standard deviation | 29 | 102 | 10 |
Table 1: Representative protoplast transfection efficiency. Data from nine recent protoplasting trials in the authors' laboratory.
Supplemental File 1: GenBank-formatted text file with the sequence of pJT01. A 4.3 kb sGFP expression plasmid serves as a positive control in transfected maize protoplasts. Please click here to download this File.
