A capacidade para projetar o genoma de qualquer organismo vivo tem muitas aplicações biomédicas e biotecnológicas, como a correção da doença-causando mutações, construção de modelos de celulares precisos para estudos de doença, ou geração de produtos agrícolas culturas com características desejáveis. Desde a virada do século, diversas tecnologias foram desenvolvidas para a engenharia do genoma em células de mamíferos, incluindo meganucleases1,2,3, zinco dedo nucleases4,5, ou transcrição como ativador effector nucleases (TALENs)6,7,8,9. No entanto, estas tecnologias anteriores são difíceis de programa ou tedioso para montar, prejudicando, assim, sua adoção generalizada na investigação e a indústria.
Nos últimos anos, o cluster regularmente intercaladas curtas palíndromos repetições (CRISPR) – sistema CRISPR-associado (Cas) tem emergido como um poderoso novo genoma engenharia tecnologia10,11. Originalmente um sistema imune adaptativo em bactérias, tem sido com sucesso implantado para a modificação do genoma em plantas e animais, incluindo seres humanos. Uma razão principal por que CRISPR-Cas ganhou tanta popularidade em tão pouco tempo é que o elemento que traz a chave endonuclease de Cas, tais como Cas9 ou Cas12a (também conhecido como Cpf1), para o local correto no genoma é simplesmente um pedaço curto de quimérico único guia RN Um (sgRNA), que é simples para o projeto e barato para sintetizar. Depois de ser recrutado para o site de destino, a enzima Cas funciona como um par de tesouras moleculares e cliva o DNA acoplado com sua RuvC, HNH ou Nuc domínios12,13,14. A resultante quebra encalhada dobro (DSB) é posteriormente reparada pelas células através de fim não-homóloga (NHEJ) juntar-se ou no caminho de reparação homologia-dirigido (HDR). Na ausência de um modelo de reparação, o ORL é reparado por via NHEJ propenso a erro, que pode dar origem a pseudo-aleatório inserção ou supressão de nucleotídeos (puntuais) no local do corte, potencialmente causando frameshift mutações nos genes codificantes de proteínas. No entanto, na presença de um modelo de doador que contém as alterações de DNA desejadas, o ORL é reparado por via HDR de alta fidelidade. Tipos comuns de modelos de doadores incluem oligonucleotides single-stranded (ssODNs) e plasmídeos. O primeiro costuma ser usado se as alterações de DNA pretendidas são pequenas (por exemplo, a alteração de um único par de base), enquanto o último é geralmente usado se deseja inserir uma sequência relativamente longa (por exemplo, a sequência de codificação de uma proteína verde fluorescente ou GFP) para o local de destino.
A atividade de endonuclease da proteína Cas requer a presença de um motivo protospacer adjacentes (PAM) para o local de destino15. O PAM de Cas9 é na extremidade 3′ da protospacer, enquanto o PAM de Cas12a (também chamado Cpf1) é, ao invés, na extremidade 5′16. O Cas-guia complexo RNA é incapaz de apresentar um ORL se o PAM estiver ausente17. Daí, o PAM coloca uma restrição na genômicas locais onde um determinado nuclease Cas é capaz de decompor. Felizmente, nucleases de Cas de diferentes espécies bacterianas geralmente apresentam exigências diferentes do PAM. Portanto, integrando vários sistemas CRISPR-Cas em nossa caixa de ferramentas de engenharia, podemos expandir a gama de sites que podem ser alvo de um genoma. Além disso, uma enzima natural do Cas pode ser projetada ou evoluiu para reconhecer sequências de PAM alternativas, ainda mais, alargar o âmbito da genômicos alvos acessíveis para manipulação de19,18,20.
Embora vários sistemas CRISPR-Cas estão disponíveis para fins de engenharia do genoma, a maioria dos usuários da tecnologia dependem principalmente da nuclease Cas9 de Streptococcus pyogenes (SpCas9) por várias razões. Em primeiro lugar, requer um PAM relativamente simplesmente NGG, ao contrário de muitas outras proteínas de Cas que podem cleave apenas na presença de PAMs mais complexas. Em segundo lugar, é a primeira endonuclease Cas seja implantado com sucesso em células humanas21,22,23,24. Em terceiro lugar, SpCas9 é, de longe, a melhor enzima caracterizada até à data. Se um pesquisador deseja usar outro nuclease Cas, ele ou ela muitas vezes seria incerto sobre qual a melhor forma de projetar o experimento e bem como outras enzimas irão realizar em diferentes contextos biológicos em relação ao SpCas9.
Para fornecer clareza para o desempenho relativo dos diferentes sistemas de CRISPR-Cas, recentemente Efetuamos uma comparação sistemática de cinco endonucleases de Cas-SpCas9, a enzima Cas9 de Staphylococcus aureus (SaCas9), a enzima Cas9 de Neisseria meningitidis (NmCas9), a enzima Cas12a de SP Acidaminococcus BV3L6 (AsCas12a) e a enzima Cas12a de bactéria Lachnospiraceae ND2006 (LbCas12a)25. Para uma comparação justa, avaliamos as nucleases de Cas diferentes usando o mesmo conjunto de sites de destino e outras condições experimentais. Os parâmetros de projeto de estudo também delineado para cada sistema CRISPR-Cas, que serviu como uma referência útil para os usuários da tecnologia. Aqui, para melhor permitir aos investigadores fazer uso das CRISPR-Cas sistema, nós fornecemos um protocolo passo a passo para engenharia de genoma ideal com diferentes enzimas Cas9 e Cas12a (ver Figura 1). O protocolo inclui não apenas detalhes experimentais, mas considerações de design também é importante para maximizar a probabilidade de um resultado de sucesso do genoma engenharia em células de mamíferos.
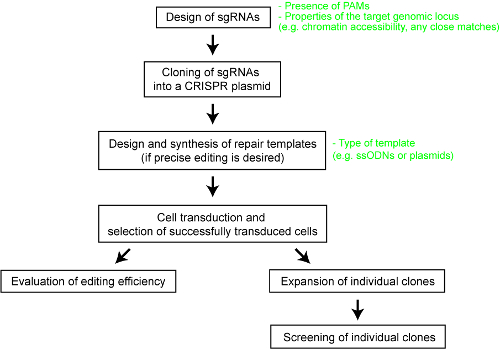
Figura 1 : Uma visão geral do fluxo de trabalho para gerar o genoma edição linha celular humana. Clique aqui para ver uma versão maior desta figura.
