الدهون هي المكونات الرئيسية للأغشية ، والتي توفر حدودا للخلايا وتمكن من التقسيم داخل الخلايا1،2،3. الدهون هي البرمائيات ، مع مجموعة رأس قطبية واثنين من ذيول الأحماض الدهنية الكارهة للماء. تتجمع هذه ذاتيا في طبقة ثنائية لتقليل ملامسة السلاسل الكارهة للماءبالماء 3,4. ينتج عن مجموعات مختلفة من مجموعات الرأس المحبة للماء والذيول الكارهة للماء فئات مختلفة من الليبيدات في الأغشية البيولوجية ، مثل الجليسيروفوسفوليبيدات ، والشحميات السفينغوبية ، والستيرول (الشكل 1) 1،5،6. الغليسيروفوسفوليبيدات هي اللبنات الأساسية لأغشية الخلايا حقيقية النواة المكونة من الجليسيروفوسفات، والأحماض الدهنية طويلة السلسلة، ومجموعات الرأس ذات الوزن الجزيئيالمنخفض 7. تعتمد تسميات الدهون على الاختلافات في مجموعات الرأس. ومن الأمثلة على ذلك فوسفاتيديل كولين (PC) ، فوسفاتيديل إيثانولامين (PE) ، فوسفاتيديل سيرين (PS) ، فوسفاتيديل جليسرول (PG) ، فوسفاتيديل إينوسيتول (PI) ، أو حمض الفوسفاتيديك غير المعدل (PA) 5,6. أما بالنسبة للذيول الكارهة للماء ، فإن طول ودرجة التشبع تختلف ، جنبا إلى جنب مع بنية العمود الفقري. التوليفات المحتملة عديدة ، مما يؤدي إلى الآلاف من أنواع الدهون في خلايا الثدييات6. تؤدي التغييرات في تكوين الدهون الغشائية إلى خصائص غشاء ميكانيكية وهيكلية مختلفة تؤثر على نشاط كل من بروتينات الغشاء المتكامل والبروتينات المحيطية 2,6.
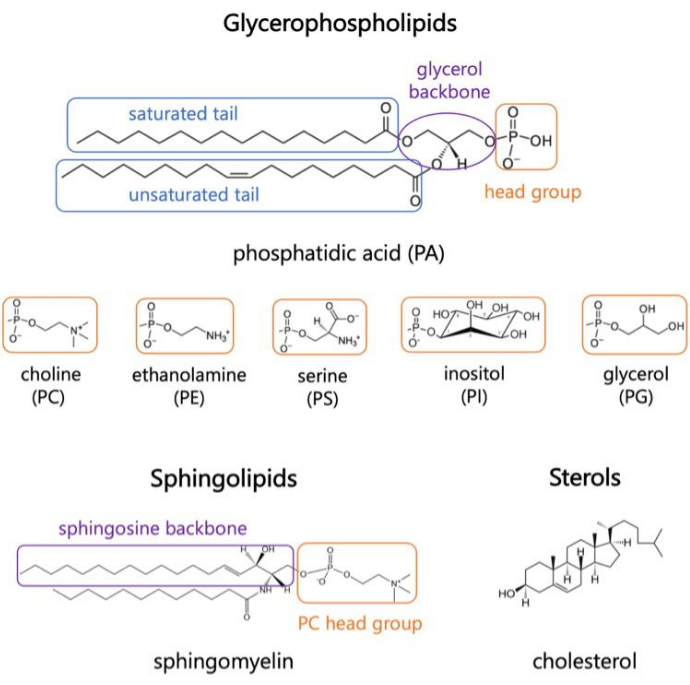
الشكل 1. هياكل الدهون التمثيلية. تظهر ذيول الأحماض الدهنية في مربعات زرقاء، ومجموعات رؤوس الليبيدات الشائعة باللون البرتقالي، والعمود الفقري للعينة باللون الأرجواني. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
الدهون هي لاعبين نشطين في العمليات الخلوية ، وتنشيط البروتين في شلالات الإشارات ، وتوازن الخلايا السليمة 8,9. ديناميات الدهون المتغيرة هي نتيجة للعدوى أو يمكن أن تكون علامات على التسبب في المرض10،11،12،13،14،15. كحواجز للخلية ، فإن دراسة الدهون الغشائية ودورها في تغلغل الجزيئات الصغيرة ذات صلة بأنظمة توصيل الأدوية وآليات تعطيل الغشاء16,17. يؤدي التنوع الكيميائي والنسب المختلفة لأنواع الدهون عبر العضيات والأنسجة والكائنات الحية إلى ديناميكيات غشاء معقدة2. لذلك من المهم الاحتفاظ بهذه الخصائص في دراسات النمذجة للطبقات المزدوجة الدهنية ، خاصة عندما يكون الهدف من الدراسة هو فحص تفاعلات الجزيئات الحيوية الأخرى مع الغشاء. تعتمد الأنواع الدهنية التي يجب مراعاتها في النموذج على الكائن الحي والمقصورة الخلوية محل الاهتمام. على سبيل المثال ، تعتبر دهون PG مهمة لنقل الإلكترون في bateria18 الضوئي ، في حين أن دهون الإينوزيتول المفسفرة (PIPs) هي لاعبين رئيسيين في ديناميكيات غشاء البلازما (PM) وشلالات الإشارات في خلايا الثدييات 19,20. داخل الخلية ، تحتوي أغشية PM والشبكة الإندوبلازمية (ER) و Golgi وأغشية الميتوكوندريا على وفرة فريدة من الدهون تؤثر على وظيفتها. على سبيل المثال ، ER هو محور التكوين الحيوي للدهون وينقل الكوليسترول إلى PM و Golgi. يحتوي على تنوع عالي الدهون مع وفرة من PC و PE ، ولكن محتوى ستيرول منخفض ، مما يعزز سيولة الغشاء21،22،23،24. في المقابل ، يشتمل PM على مئات وحتى آلاف الأنواع الدهنية اعتمادا على الكائن الحي25 ، ويحتوي على مستويات عالية من الدهون السفينغولية والكوليسترول التي تمنحه صلابة مميزة مقارنة بالأغشية الأخرى في الخلية24. يجب مراعاة عدم تناسق الوريقات للأغشية مثل PM ، التي تحتوي على نشرة خارجية غنية بالسفينغوميلين ، و PC ، والكوليسترول ، ونشرة داخلية غنية ب PE و PI و PS التي تعتبر مهمة للإشارة إلى الشلالات24. أخيرا ، يدفع تنوع الدهون أيضا إلى تكوين مجالات دقيقة تختلف في التعبئة والنظام الداخلي ، والمعروفة باسم طوافات الدهون24,26 ؛ تظهر هذه عدم التماثل الجانبي ، ويفترض أنها تلعب أدوارا مهمة في الإشارات الخلوية26 ، ويصعب دراستها بسبب طبيعتها العابرة.
تم استخدام التقنيات التجريبية مثل التنظير الفلوري والتحليل الطيفي وأنظمة الأغشية النموذجية مثل الحويصلات أحادية الصفيحة العملاقة (GUVs) للتحقيق في تفاعلات الجزيئات الحيوية مع الأغشية. ومع ذلك ، من الصعب التقاط الطبيعة المعقدة والديناميكية للمكونات المعنية بالطرق التجريبية وحدها. على سبيل المثال ، هناك قيود على تصوير المجالات عبر الغشاء للبروتينات ، وتعقيد الأغشية المستخدمة في مثل هذه الدراسات ، وتحديد الحالات الوسيطة أو العابرة أثناء عملية الاهتمام27،28،29. منذ ظهور المحاكاة الجزيئية للطبقات الأحادية الدهنية والطبقات الثنائية في ثمانينيات القرن العشرين29 ، يمكن الآن تحديد أنظمة البروتين الدهني وتفاعلاتها على المستوى الجزيئي. محاكاة الديناميات الجزيئية (MD) هي تقنية حسابية شائعة تتنبأ بحركة الجسيمات بناء على قواها بين الجزيئات. يصف جهد التفاعل الإضافي التفاعلات المرتبطة وغير المرتبطة بين جسيمات النظام30. تسمى مجموعة المعلمات المستخدمة لنمذجة هذه التفاعلات حقل قوة المحاكاة (FF). يتم الحصول على هذه المعلمات من حسابات ab initio ، والحسابات شبه التجريبية ، وميكانيكا الكم ، والأمثل للبيانات المستنسخة من تجارب الأشعة السينية وحيود الإلكترون ، والرنين المغناطيسي النووي ، والأشعة تحت الحمراء ، والتحليل الطيفي للنيوترونات ، من بين طرق أخرى31.
يمكن استخدام محاكاة MD لدراسة الأنظمة على مستويات مختلفة من الدقة32،33،34. تتم دراسة الأنظمة التي تهدف إلى توصيف تفاعلات جزيئية حيوية محددة ، وروابط هيدروجينية ، وتفاصيل أخرى عالية الدقة باستخدام محاكاة جميع الذرات (AA). في المقابل ، تقوم عمليات المحاكاة الخشنة (CG) بتجميع الذرات في مجموعات وظيفية أكبر لتقليل التكلفة الحسابية وفحص ديناميكيات النطاقالأكبر 33. تقع بين هذين المحاكاة للذرة المتحدة (UA) ، حيث يتم دمج ذرات الهيدروجين مع ذراتها الثقيلة لتسريع الحساب33,35. تعد محاكاة MD أداة قوية لاستكشاف ديناميكيات الأغشية الدهنية وتفاعلاتها مع الجزيئات الأخرى ويمكن أن تعمل على توفير آليات المستوى الجزيئي للعمليات ذات الأهمية في واجهة الغشاء. بالإضافة إلى ذلك ، يمكن أن تعمل محاكاة MD على تضييق نطاق الأهداف التجريبية والتنبؤ بالخصائص الجزيئية الكبيرة لنظام معين بناء على التفاعلات المجهرية.
باختصار ، بالنظر إلى مجموعة من الإحداثيات الأولية والسرعات ومجموعة من الشروط مثل درجة الحرارة والضغط الثابتين ، يتم حساب مواضع وسرعات كل جسيم من خلال التكامل العددي لجهد التفاعل وقانون نيوتن للحركة. يتكرر هذا بشكل متكرر ، وبالتالي توليد مسار محاكاة30. يتم إجراء هذه الحسابات باستخدام محرك MD ؛ من بين العديد من الحزم مفتوحة المصدر ، يعد GROMACS36 أحد أكثر المحركات استخداما والذي وصفناه هنا. ويشمل أيضا أدوات لتحليل وبناء الإحداثيات الأولية للنظم المراد محاكاتها37. تشمل محركات MD الأخرى NAMD38. CHARMM39 و AMBER40 ، والتي يمكن للمستخدم اختيارها وفقا لتقديره الخاص بناء على الأداء الحسابي لنظام معين. من الأهمية بمكان تصور المسارات أثناء المحاكاة وكذلك لتحليل النتائج وتفسيرها. تتوفر مجموعة متنوعة من الأدوات. نناقش هنا الديناميات الجزيئية المرئية (VMD) التي تقدم مجموعة واسعة من الميزات ، بما في ذلك التصور ثلاثي الأبعاد (3-D) مع طرق الرسم والتلوين الموسعة ، وتصور البيانات الحجمي ، وبناء وإعداد وتحليل مسارات أنظمة محاكاة MD ، وصنع فيلم المسار بدون حدود لحجم النظام ، إذا كانت الذاكرة متوفرة41،42،43.
تتأثر دقة الديناميكيات المتوقعة بين مكونات النظام بشكل مباشر ب FF المختار لنشر المسار. يتم متابعة جهود معلمات FF التجريبية من قبل عدد قليل من مجموعات البحث. تشمل FF الأكثر رسوخا وشيوعا ل MD CHARMM39 و AMBER 40 و Martini44 و OPLS 45 و SIRAH 46. يستخدم مجال القوة المضافة لجميع الذرات CHARMM36 (C36)47 على نطاق واسع ل AA MD لأنظمة الأغشية لأنه يعيد إنتاج البيانات الهيكلية التجريبية بدقة. تم تطويره في الأصل من قبل مجتمع CHARMM ، وهو متوافق مع العديد من محركات MD مثل GROMACS و NAMD. على الرغم من التحسينات عبر FFs الشائعة ، هناك جهد مستمر لتحسين مجموعات المعلمات للسماح بالتنبؤات التي تعيد إنتاج الملاحظات التجريبية عن كثب ، مدفوعة بالمصالح في أنظمة معينة للدراسة48,49.
يتمثل التحدي عند محاكاة الأغشية الدهنية في تحديد طول مسار المحاكاة. يعتمد هذا إلى حد كبير على المقاييس التي سيتم تحليلها والعملية التي يهدف المرء إلى توصيفها. عادة ما تتطلب مخاليط الليبيدات المعقدة وقتا أطول للوصول إلى الاتزان؛ حيث يجب أن يكون لدى المزيد من الأنواع وقت كاف للانتشار على مستوى الغشاء والوصول إلى تنظيم جانبي مستقر. يقال إن المحاكاة تكون في حالة توازن عندما تصل خاصية الاهتمام إلى هضبة وتتقلب حول قيمة ثابتة. من الممارسات الشائعة الحصول على ما لا يقل عن 100-200 نانوثانية من المسار المتوازن لإجراء تحليل إحصائي مناسب على الخصائص والتفاعلات ذات الاهتمام. من الشائع إجراء عمليات محاكاة غشائية فقط بين 200-500 نانوثانية ، اعتمادا على مدى تعقيد خليط الدهون وسؤال البحث. تتطلب تفاعلات البروتين والدهون عادة أوقات محاكاة أطول ، بين 500-2000 نانوثانية. وفيما يلي بعض النهج لتسريع أخذ العينات والديناميات التي يمكن ملاحظتها باستخدام النظم الغشائية: ‘1’ نموذج المحاكاة الغشائية العالية الحركة، الذي يستعيض عن الكربونات الطرفية للدهون في الغشاء بالمذيب العضوي لتسريع أخذ العينات 50؛ و ‘2’ نموذج المحاكاة الغشائية العالية الحركة، الذي يستعيض عن الكربونات الطرفية للدهون في الغشاء بالمذيب العضوي لتسريع أخذ العينات 50؛ و ‘2’ نموذج المحاكاة الغشائية العالية الحركة، الذي يستعيض عن الكربونات الطرفية للدهون في الغشاء بالمذيب العضوي لتسريع أخذ العينات50؛ و ‘2’ نموذج المحاكاة الغشائية العالية الحركة، الذي يستعيض عن الكربونات الغشائية العالية الحركة (HMMM)، الذي يستعيض عن و (ii) إعادة تقسيم كتلة الهيدروجين (HMR) ، والذي يجمع جزءا صغيرا من كتل الذرات الثقيلة داخل النظام مع تلك الخاصة بذرات الهيدروجين للسماح باستخدام محاكاة أكبر للخطوةالزمنية 51.
يناقش البروتوكول التالي نهجا صديقا للمبتدئين لبناء وتشغيل وتحليل نماذج الأغشية الواقعية باستخدام AA MD. نظرا لطبيعة محاكاة MD ، يجب تشغيل مسارات متعددة لمراعاة قابلية التكرار والتحليل الإحصائي المناسب للنتائج. من الممارسات الحالية تشغيل ثلاث نسخ متماثلة على الأقل لكل نظام من أنظمة الاهتمام. بمجرد اختيار الأنواع الليبيدة للكائن الحي والعملية محل الاهتمام، يتم توضيح وتلخيص الخطوات الأساسية لبناء مسار محاكاة لنظام الغشاء فقط وتشغيله وتحليله.
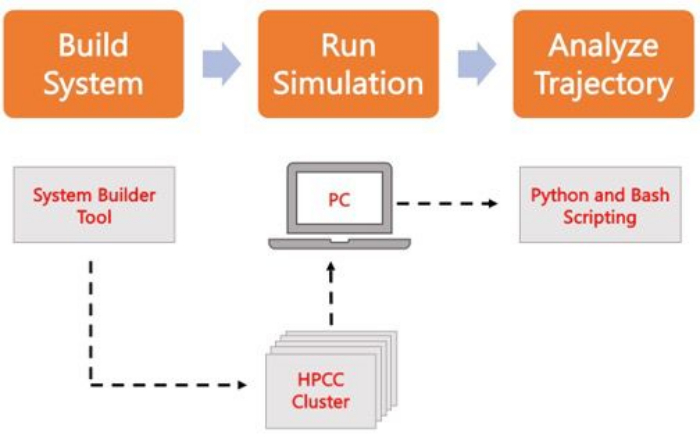
الشكل 2. تخطيطي لتشغيل محاكاة MD. تتوافق المربعات البرتقالية مع الخطوات الرئيسية الثلاث الموضحة في البروتوكول. يوجد تحتها سير عمل عملية المحاكاة. أثناء إعداد النظام ، يتم بناء النظام الذي يحتوي على الإحداثيات الأولية لنظام الغشاء المذاب باستخدام مولد إدخال النظام مثل CHARMM-GUI Membrane Builder. بعد نقل ملفات الإدخال إلى مجموعة حوسبة عالية الأداء ، يتم نشر مسار المحاكاة باستخدام محرك MD ، مثل GROMACS. يمكن إجراء تحليل المسار على مجموعة الكمبيوتر أو محطة عمل محلية جنبا إلى جنب مع التصور. ثم يتم إجراء التحليل ، باستخدام الحزم التي تحتوي على كود تحليل مدمج مثل GROMACS و VMD ، أو باستخدام نصوص Bash أو مكتبات Python المختلفة. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
