The presented method can be reliably used to record Vm in cannulated vessels. A brief procedure describing how to isolate MCA from the brain is presented in Figure 1A. After separating the brain from the skull, the MCA was dissected out and placed in a Petri dish containing low calcium PSS. Part of the connective tissue that was attached was also dissected along with MCA using spring scissors and forceps to prevent damage to MCA during the isolation. Carefully, connective tissue was also removed, and the dissected MCA was ready to transfer to the myograph. MCA was mounted on the cannulas and tied using suture knots on both ends of the cannulas in the myograph. A schematic representation of a typical microelectrode impalement method setup is shown in Figure 1B. A microelectrode filled with 3 M KCl was connected to the electrometer via a holder in the probe. Channel output of the electrometer was connected to an analog input channel of a digitizer using a BNC-BNC cable. Digitizer output was further connected to an oscilloscope to visualize the signal in the recording software. The ground is established using an AgCl pellet wire that was extended from the chassis of electrometer to the bath solution in the myograph. Finally, digital traces were visualized in the recording software on the computer monitor.
The MCA was then incubated in freshly prepared warm PSS and was pressurized to 60 mmHg. Only vessels that gain tone were used for Vm recording. After the vessel gained significant tone, the microelectrode was advanced into the vessel wall. Arterial diameter and impalement of the artery were visualized using video microscopy. Impalement is considered successful when there is a rapid deflection to negative values, Vm is stable for ≥30 s, and the voltage returns abruptly to 0 mV upon removal of the electrode as shown in Figure 212,15. Our results suggest that, in an MCA that is pressurized to 60 mmHg, the Vm is ~ -43.2 ± 2.9 mV.
After a successful impalement and Vm stabilization, the drugs that change the Vm were perfused in the bath and changes in the Vm were recorded. We used 20 mM KCl to depolarize and 20 µM NS1619, a synthetic large conductance potassium channel opener, to hyperpolarize the membrane. Our results suggest that perfusion of the chamber with KCl depolarized the membrane by ~ 5.8 ± 0.18 mV. On the other hand, perfusion with NS1619 hyperpolarized the membrane by ~3.8 ± 0.4 mV.
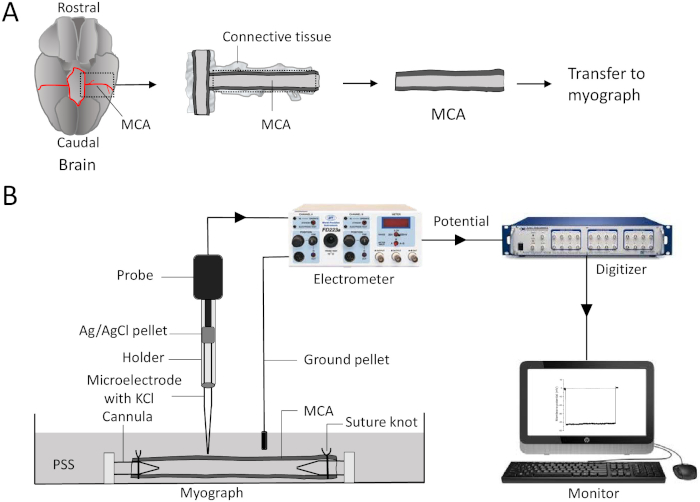
Figure 1: An illustration of isolation of middle cerebral arteries and recording of membrane potential using microelectrode impalement method. (A) Using spring scissors, cut the brain or connective tissue along the dotted lines. Transfer and mount the MCA between the two glass cannulas and secure it using sutures in the myograph chamber filled with PSS. (B) A microelectrode filled with 3 M KCl is inserted into the holder and is connected to the probe. Changes in transmembrane potential travel from the probe to an electrometer and to the digitizer via a BNC cable (BNC cable is connected from channel output of electrometer and to an analog input of a digitizer). The digitized potential is seen in the recording software on the monitor. The ground is established using an AgCl pellet wire connected from the electrometer to the bath solution in the myograph. MCA = middle cerebral artery; PSS = physiological salt solution; AgCl = silver chloride. Please click here to view a larger version of this figure.
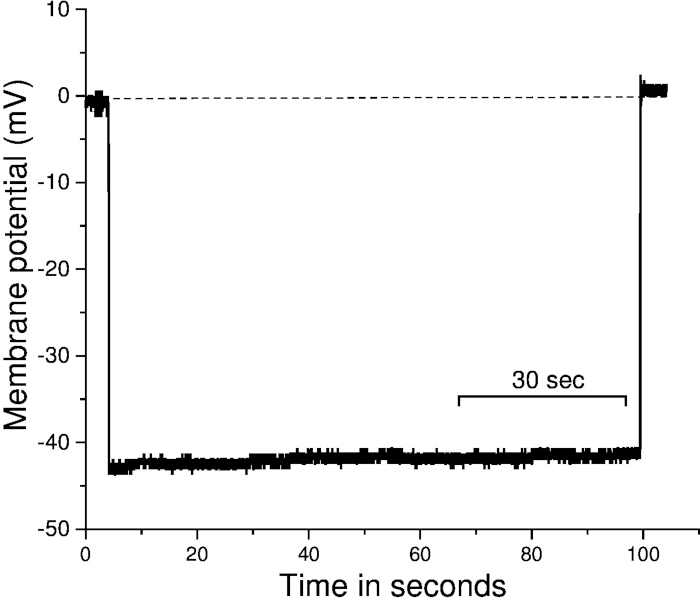
Figure 2: Recording of membrane potential using microelectrode impalement method from cannulated middle cerebral arteries. Representative trace of Vm. Impalement is considered successful if there is an abrupt deflection to negative values upon electrode entry, Vm is stable for ≥30 s, and the voltage returns abruptly to 0 mV upon removal of the electrode. The X-axis represent the time, and the Y-axis represents the membrane potential. The dotted line represents the adjusted baseline before the impalement of the vessel. Sec = seconds. Please click here to view a larger version of this figure.
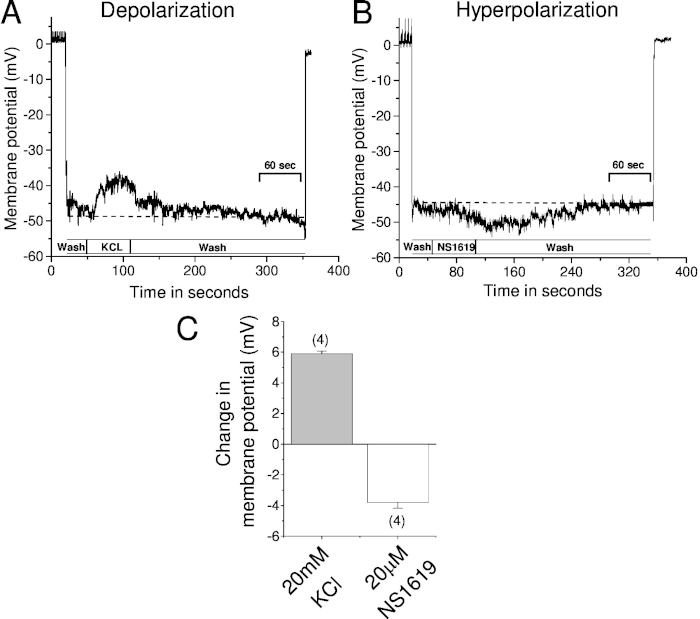
Figure 3: Recording of changes in the membrane potential using microelectrode impalement method when vessels are exposed to vasoactive agents. Vm was recorded using microelectrode impalement method from cannulated middle cerebral arteries before and after exposure to vasoactive agents. Sample trace represents (A) depolarization in response to 20 mM KCl and (B) hyperpolarization in response to 20 µM NS1619—a large conductance potassium channel agonist. (C) Summary bar graph of the changes in Vm before and after application of KCl and NS1619. The dotted line represents resting Vm. The number in the parenthesis represents the number of vessels used in the study. Note that Vm reached baseline as soon as the drug is washed out. Error bar represents the standard error of the mean. Sec = seconds. Please click here to view a larger version of this figure.
| Settings of electrometer to measure electrode resistance | |
| Meter Input | Channel A or B |
| Position toggle | In |
| Meter range | toggle to 200 mV |
| Electrode | In the bath |
| Mode | Operate to Electrode test |
| Meter indication | 1 mV/MΩ |
Table 1: Settings of electrometer to measure electrode resistance.
| Chemicals | low calcium PSS mM | Normal PSS mM | Company | Catalog | |
| 1 | NaCl | 119.00 | 119.00 | Sigma | S7653 |
| 2 | KCl | 4.70 | 4.70 | Sigma | P4504 |
| 3 | MgSO4 | 1.17 | 1.17 | Sigma | M7506 |
| 4 | CaCl2 | 0.05 | 1.60 | Sigma | C3881 |
| 5 | HEPES | 5.00 | 5.00 | Sigma | H7006 |
| 6 | Glucose | 10.00 | 10.00 | Sigma | G7021 |
| 7 | NaH2PO4 | 1.18 | 1.18 | Sigma | S0751 |
| 8 | NaHCO3 | 18.00 | 18.00 | Sigma | S5761 |
| pH: 7.4 | pH: 7.4 |
Table 2: Reagents used in the preparation of low calcium and normal physiological salt solution.
