Wnt/β-catenin activity
Whole mount Lac-Z staining was detected in tracheal-lung tissue of embryos isolated from reporter Axin2Lac-Z mice11. Sites of staining indicate Wnt/β-catenin activity. Analysis of sections of whole mount staining determined that Wnt/β-catenin activity was present in the mesenchyme of the trachea and in mesenchyme of peripheral regions of developing lungs. In WlsShhCre embryos (wherein secretion of Wnt ligands from the epithelium of respiratory tract was abrogated) Lac-Z staining was almost absent (Figure 1).
Mesenchymal condensations in tracheal mesenchyme
A critical step in chondrogenesis is the process by which mesenchymal cells fated to give rise to cartilage condense. These tight aggregations of cells are known as mesenchymal condensations. To test whether the lack of cartilage observed in WlsShhCre mice was due to an absence of mesenchymal condensations, tracheal lung tissues from gestational ages E12.5 to E14.5 were stained with PNA lectin. At E13.5 and E14.5, fluorescence was detected as periodic bands in the tracheal mesenchyme at sites where cartilage will be formed. Lack of detectable fluorescence in the tracheal tissue of WlsShhCre embryos indicates that endodermal Wnt signaling to the tracheal mesenchyme is necessary for mesenchymal condensations (Figure 2).
Respiratory tract cell specification
Whole mount immunofluorescence of tracheal lung tissues determined the expression pattern of Sox9, Nkx2.1 and αSMA at E11.5. In the tracheal tissue, Sox9 expression is limited to the mesenchyme of the trachea as a continuous stripe (arrow in Figure 3A and B) while in the developing lung Sox9 is observed at the periphery of the respiratory epithelium (arrow heads in Figure 3A, B and C). Nkx2.1 presents a distinct expression pattern restricted to the epithelium of the trachea of the lung. Preventing secretion of Wnt ligands from epithelium causes diminished expression of Sox9 in tracheal mesenchyme but does not affect Sox9 expression in peripheral lung epithelium. Expression of αSMA is not readily detected in tracheal mesenchyme, however staining was detected at the level of the larynx (yellow arrow, Figure 3) and stomach (Figure 3).
Cell proliferation in developing trachea
Proliferation in epithelium and mesenchyme of the trachea was detected after in vivo labeling of tissue with BrdU. Sections of tracheal tissue were stained with antibodies recognizing BrdU, Sox9 and αSMA. The proliferation profile in the tracheal mesenchyme indicates that the levels of Sox9 stained cells undergoing proliferation is higher than the level of αSMA stained cells undergoing proliferation. This proliferation pattern seems to be reverted in WlsShhCre tracheas (Figure 4).
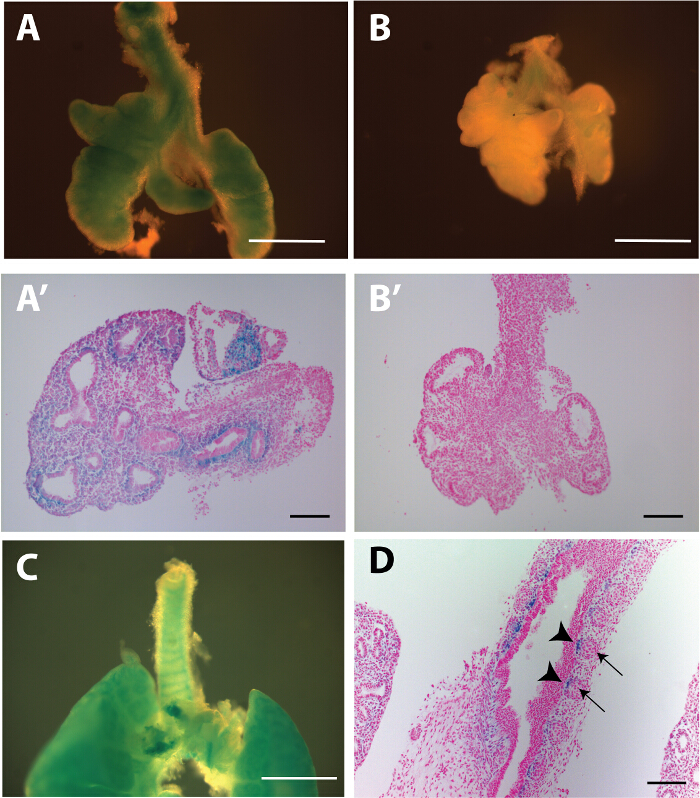
Figure 1: Wnt/β-catenin activity is operative in tracheal mesenchyme. E13.5 tracheal lung tissue, isolated from Axin2-LacZ and WlsShhCre:Axin2-LacZ mice, were stained with X-gal (A and B). Sections of tracheal lung tissue were counterstained with fast red (A', B'). Note the lack of X-gal staining in WlsShhCre;Axin2-Lac-Z mice demonstrating absence of Wnt/β-catenin activity (B'). E14.5 tissue from Axin2 LacZ mice stained with X-gal depicts sites of future cartilaginous rings (C); however, Axin2 activity was restricted to the periphery (arrow head) of the mesenchymal condensations (arrow) (D). Scale bar A, B and C = 500 μm; B', D' and D = 100 μm. Please click here to view a larger version of this figure.
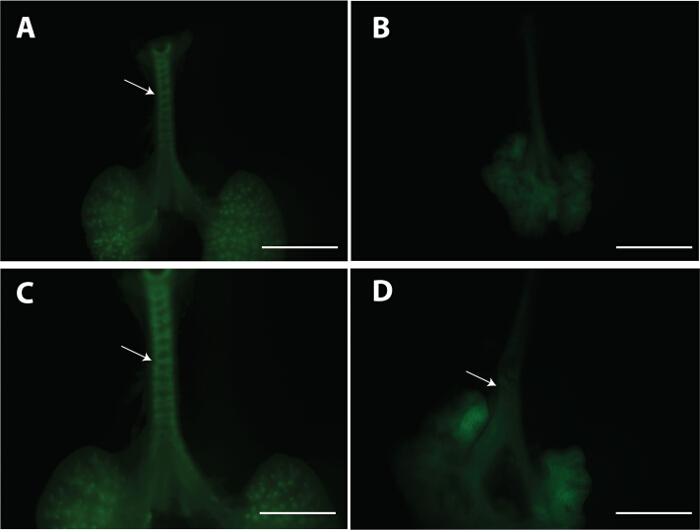
Figure 2: Mesenchymal condensation in developing tracheal tissue. E13.5 tracheal-lung explants were stained with PNA lectin. Control tracheal tissue depicting the regions were mesenchymal cells condensed is shown (arrows A, C). No mesenchymal condensations were detected in WlsShhCre mice (B and arrow in D). C and D depict lower magnification of tracheal-lung tissue shown in A and B. Scale bar A and B = 1 mm; C and D = 2 mm. Please click here to view a larger version of this figure.
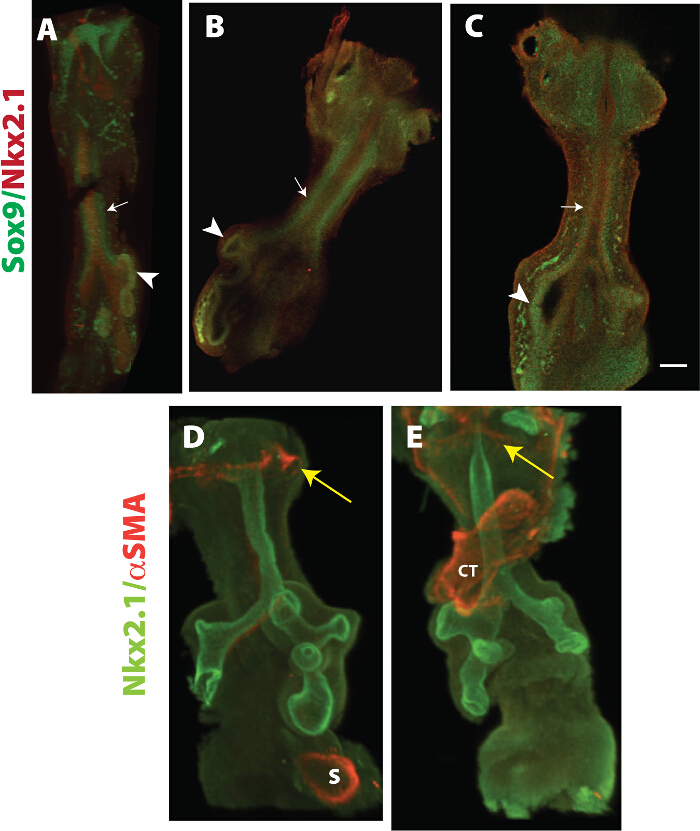
Figure 3: Expression pattern in prechondrogenic whole mount trachea. Nkx2.1 and Sox9 staining was detected in E11.5 explants. Panel A depicts an image of a control tissue, while panel B shows an optical section of a 20 μm control tissue. Note the lack of mesenchymal expression of Sox9 in WlsShhCre tissue (arrow in C), but maintenance of peripheral expression of Sox9 (arrowhead in C). αSMA staining was detected at low levels in trachealmesenchyme of E11.5 tissue but observed in the laryngeal region (yellow arrowhead in D and E), stomach (S) and remaining cardiac tissue (CT) (E). Nkx2.1 is expressed in epithelium of developing trachea and lungs (D, E). Scale bar = 10 µm. Please click here to view a larger version of this figure.
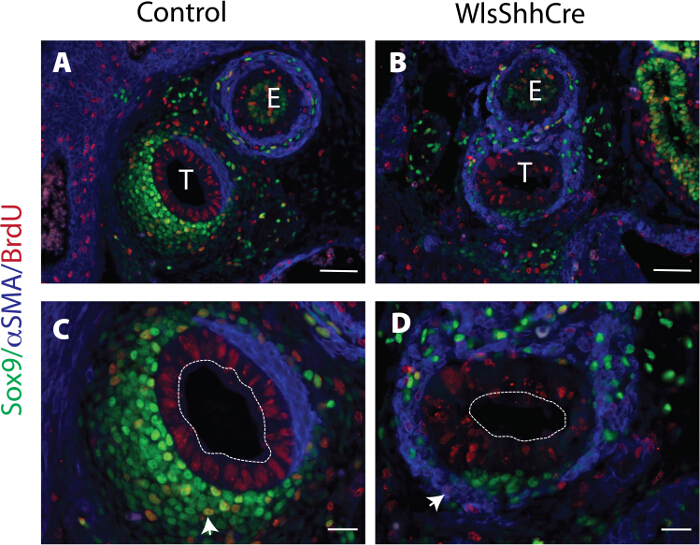
Figure 4: Cell proliferation pattern in prechondrogenic mesenchyme. Sections of E13.5 embryos were stained with anti BrdU, Sox9 and αSMA antibody. Panels C and D are higher magnifications of A and B respectively. Dotted lines represent limits of the tracheal epithelium. Note Sox9 stained proliferative cell (arrowhead C) and αSMA stained proliferative cell (arrowhead in D). T = trachea, E = esophagus. Scale bar A and B = 50 μm, C and D = 20 μm. Please click here to view a larger version of this figure.
