التفاعل بين الجزيئات هو أساس الطبيعة. ومن هنا، والعلماء في العديد من مجالات البحوث الأساسية والتطبيقية محاولة لفهم المبادئ الأساسية للتفاعلات الجزيئية من أنواع مختلفة. الميكروسكيل Thermophoresis (MST) تمكن العلماء من أداء ودقيقة، وتوصيف سريع، ومراقبة الجودة فعالة من حيث التكلفة من التفاعلات الجزيئية في الحل، مع حرية اختيار المخازن. وهناك بالفعل أكثر من 1000 المنشورات باستخدام MST، اعتبارا من عام 2016 وحده، واصفا أنواع مختلفة من التحليلات، بما في ذلك عروض مكتبة، ملزمة التصديقات الحدث، فحوصات المنافسة، والتجارب مع شركاء ملزم متعددة 1-8. بشكل عام، يسمح MST دراسة المعلمات ملزمة الكلاسيكية، مثل تقارب ملزمة (بعد الظهر لملم)، رياضيات الكيمياء، والديناميكا الحرارية، من أي نوع من التفاعل الجزيئي. وهناك ميزة كبيرة من MST هي القدرة على دراسة الأحداث ملزمة مستقلة عن حجم شركاء التفاعل. حتى شالالتفاعلات lenging بين الأبتامرات الحمض النووي صغيرة (15-30 الإقليم الشمالي) وأهداف مثل الجزيئات الصغيرة، والمخدرات، والمضادات الحيوية، أو الأيض ويمكن قياس.
التكنولوجيات الحالية للدولة من بين الفن لتوصيف التفاعلات أبتمر المستهدفة هي إما مختبر مكثفة ومعقدة للغاية أو تفشل في تحديد-أبتمر صغيرة جزيء التفاعلات 9،10. السطح بلازمون الرنين (SPR) المستندة إلى فحوصات 11،12 والنهج المسعرية حقا خالية من التسمية، مثل متساوي الكالوري المعايرة (ITC) 13-15، شطف isocratic 16، التوازن فاي ltration 17،18، في خط التحقيق 19، gel- تحول المقايسات، stopped- آه فلوريدا فلوريدا uorescence الطيفي 20،21، مضان تباين (FA) 22،23، واحد جزيء فلوريدا uorescence التصوير 24،25، والحيوية طبقة التداخل (BLI) 26 هي أيضا إما غير دقيق أو غير متوافق مع جزيء صغير أبتمر التفاعلات. principa الآخرينقضايا لتر من هذه الأساليب هي حساسية منخفضة، وارتفاع استهلاك عينة، الشلل، والقيود النقل الجماعي على الأسطح، و / أو قيود العازلة. فقط عدد قليل من هذه التقنيات توفر ضوابط متكاملة لتجميع وامتصاص الآثار.
يمثل MST أداة قوية للعلماء للتغلب على هذا القيد لدراسة التفاعلات بين الأبتامرات والجزيئات الصغيرة 27-29، فضلا عن أهداف أخرى مثل البروتينات 30-33. وتعتمد هذه التقنية على حركة الجزيئات من خلال تدرجات درجة الحرارة. هذه الحركة موجهة، ودعا "thermophoresis،" تعتمد على حجم وتوجيه التهم وقذيفة الماء من جزيء 34،35. الربط ليجند للجزيء ستغير مباشرة واحدة على الأقل من هذه المعايير، مما أدى إلى التنقل thermophoretic تغييرها. بروابط مع أحجام صغيرة قد لا يكون لها تأثير كبير من حيث حجم التغيير من غير منضم إلى دولة منضمة، ولكنها يمكن أن يكون الدكتور آثار amatic على قذيفة الماء و / أو تهمة. التغييرات في حركة thermophoretic من جزيئات بعد التفاعل مع الشريك ملزم تمكن الكمي من المعلمات ملزمة الأساسية 2،7،34،36،37.
كما هو مبين في الشكل 1A، ويتكون الجهاز MST ليزر الأشعة تحت الحمراء التي تركز على العينة داخل الشعيرات الدموية الزجاج باستخدام نفس البصريات، وللكشف عن مضان. يمكن رصد حركة thermophoretic من البروتينات عن طريق uorescence فلوريدا لا يتجزأ من tryptophans 6 أو تفاعل شريك 3،8 fluorescently المسمى في حين أن الليزر يؤسس التدرج في درجة الحرارة (ΔT من 2-6 درجة مئوية). الفرق في درجة الحرارة مما أدى إلى الفضاء، ΔT، يؤدي إلى نضوب أو تراكم الجزيئات في مجال درجات الحرارة المرتفعة، والتي يمكن قياسها كميا بواسطة سوريه coef فاي cient (S T):
ز "/>
ج يمثل الساخنة التركيز في منطقة ساخنة، وج البرد هو تركيز في المنطقة الباردة الأولية.
كما هو مبين في الشكل 1B، نموذجية MST نتائج التجربة في ملف تعريف حركة MST (الوقت أثر)، ويتألف من المراحل المختلفة، والتي يمكن أن تكون مفصولة الجداول الزمنية الخاصة بكل منها. يتم قياس مضان الأولي في أول 5 ثوان في غياب التدرج في درجة الحرارة لتحديد مضان بدءا دقيقة وللتحقق ل photobleaching أو photoenhancement. الوثب درجة الحرارة (T-السريع) يمثل المرحلة التي التغييرات مضان قبل حركة thermophoretic. هذا الانخفاض الأولي في مضان يعتمد على التغييرات التي تعتمد على الحرارة من فلوريدا uorophore العائد الكم. يتبع مرحلة thermophoresis، التي ينخفض مضان (أو زيادة) نتيجة لحركة thermophoretic من الجزيئات حتى توزيع ثابتة للدولة يتم التوصل.ويمكن ملاحظة أن TJump العكس ونشر الظهر يصاحب ذلك من الجزيئات uorescent فلوريدا كما هو مبين في الشكل 1B بعد يتم فيها تشغيل الليزر قبالة. من أجل الوصول إلى معايير ملزمة الأساسية، ويتم تحليل نسب المولي مختلفة من شركاء التفاعل ومقارنة. عادة، يتم دراسة 16 نسب مختلفة في تجربة MST واحد، في حين يتم الاحتفاظ جزيء مرئية بصري مستمر ويتم توفيره مع كمية متزايدة من يجند الخالي من الملصقات. التفاعل بين الشركاء ملزم اثنين يؤدي الى تغييرات في thermophoresis، وبالتالي في uorescence فلوريدا تطبيع، F القاعدة، والذي يحسب على النحو التالي:
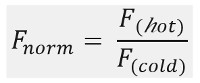
F الساخنة والباردة F تمثل متوسط كثافة uorescence فلوريدا في دي فاي نقطة زمنية نيد من آثار MST. الانتماءات الملزمة (ك د أو EC 50 القيم) ويمكن حساب من المنحنياتالبريد المناسب (الشكل 1C).
وعموما، MST هو أداة قوية لدراسة التفاعلات الجزيئية من أي نوع. تقدم هذه المخطوطة بروتوكول لوصف التفاعل تحديا بين ثلاثي صغيرة جزيء الفوسفات (ATP؛ 0.5 كيلو دالتون) و25 الإقليم الشمالي القصير ssDNA أبتمر DH25.42 (7.9 كيلو دالتون). على مدى المخطوطة، يتم تعيين موقع ملزم للأبتمر على جزيء ATP وصولا الى مجموعة الأدينين من لاعبي التنس المحترفين.
