Pseudomonas aeruginosa (Pa) è un patogeno opportunistico che stabilisce infezioni croniche in individui immuno-compromessi. Per quelli con la malattia genetica fibrosi cistica (FC), queste infezioni possono durare nel corso della vita. La FC provoca l’accumulo di un espettorato viscoso e ricco di sostanze nutritive nelle vie aeree, che viene colonizzato da una varietà di agenti patogeni microbici nel tempo. Pa è uno dei patogeni CF più diffusi, colonizzando le vie aeree nella prima infanzia e stabilendo infezioni difficili da trattare1. La Pa rimane un problema clinico significativo ed è considerata una delle principali cause di mortalità in quelli con FC, nonostante i regimi terapeutici migliorati negli ultimi anni2,3. Questo fenotipo di persistenza e la crescente tolleranza agli antibiotici hanno fatto guadagnare a Pa un posto in un gruppo di patogeni identificati sia dai Centers for Disease Control (CDC) che dall’Organizzazione Mondiale della Sanità (OMS) come priorità di ricerca per lo sviluppo di nuove strategie terapeutiche: i patogeni ESKAPE4.
Come altri patogeni ESKAPE, la resistenza agli antibiotici acquisita è comune in Pa, ma ci sono anche molte proprietà intrinseche che contribuiscono alla tolleranza antimicrobica Pa. Tra questi c’è la capacità di Pa di formare aggregati-cluster altamente densi di ~ 10-1.000 cellule, che possono essere osservati in infezioni multiple, tra cui l’espettorato del paziente CF5,6. Simile alla Pa studiata in altri sistemi di biofilm, gli aggregati di Pa mostrano fenotipi clinicamente rilevanti come l’aumento della resistenza agli antibiotici e l’attivazione della comunicazione cellula-cellula (quorum sensing (QS)). Ad esempio, gli aggregati di Pa hanno dimostrato di utilizzare comportamenti regolati da QS per combattere altri microbi e tollerare trattamenti antimicrobici come la produzione di piocianina7. La capacità di studiare tali comportamenti offre una visione emozionante degli ecosistemi batterici in un ambiente simile a quello in cui esistono nel corpo umano.
Una delle maggiori sfide per studiare come gli aggregati di Pa rispondono al mutevole ambiente dell’espettorato è la mancanza di sistemi nutrizionalmente rilevanti e robusti che promuovano la formazione di aggregati. Gran parte di ciò che si sa sulla Pa è stato scoperto utilizzando sistemi in vitro in cui le cellule crescono planctonicamente o in una caratteristica architettura “a fungo” attaccata alla superficie che non è stata osservata in vivo8. Mentre i modelli classici di crescita del biofilm, come le cellule di flusso o l’agar solido, hanno prodotto una conoscenza ampia e preziosa sui comportamenti batterici e sui meccanismi di tolleranza agli antibiotici, questi risultati non sempre si traducono in vivo. Molti modelli in vitro hanno una capacità limitata di imitare l’ambiente di crescita del sito di infezione umana, rendendo necessari costosi studi in vivo. A loro volta, molti modelli in vivo mancano della flessibilità e della risoluzione offerte dalle tecniche in vitro.
L’espettorato sintetico della fibrosi cistica (SCFM2) è progettato per fornire un ambiente per la crescita di Pa simile a quello sperimentato durante l’infezione cronica nel polmone CF. SCFM2 include fonti nutrizionali identificate nella CF sputa espettorata oltre a mucina, lipidi e DNA. La crescita di Pa in SCFM2 richiede un set genetico quasi identico a quello richiesto per la crescita nell’espettorato reale e supporta la formazione naturale di aggregati di Pa 9,10. Dopo l’inoculazione, le cellule planctoniche formano aggregati che aumentano di dimensioni attraverso l’espansione. Le singole cellule (denominate migranti) vengono rilasciate dagli aggregati, migrano verso aree non colonizzate e formano nuovi aggregati10. Questa storia di vita può essere osservata utilizzando CLSM e l’analisi delle immagini alla risoluzione di una singola cellula. Gli aggregati di Pa formati in SCFM2 sono di dimensioni simili a quelli osservati nel polmone CF10. Questo modello consente l’osservazione di aggregati multipli di varie dimensioni in tempo reale e in tre dimensioni su scala micron. La microscopia time-lapse consente il tracciamento di migliaia (~ 50.000) di aggregati in un unico esperimento. L’uso di software di analisi delle immagini consente la quantificazione di fenotipi aggregati da micrografie, tra cui volume aggregato, superficie e posizione in tre dimensioni fino all’aeroporto di 0,1 μm, sia a livello di aggregato individuale che di popolazione. Avere la capacità di raggruppare gli aggregati per fenotipo e posizione consente la differenziazione degli aggregati in diversi stadi di sviluppo con precisione, nonché la loro risposta a un microambiente mutevole6,11.
L’applicazione di SCFM2 per studiare aggregati di Pa in saggi a basso volume e ad alto rendimento lo rendono un modello flessibile ed economico. Come mezzo definito, SCFM2 offre uniformità e riproducibilità su più piattaforme, fornendo un metodo nutrizionalmente e fisicamente rilevante per studiare gli aggregati di Pa in vitro9. Le applicazioni includono il suo uso in combinazione con CLSM per osservare l’organizzazione spaziale e la tolleranza agli antibiotici ad alta risoluzione (come descritto in questo documento sui metodi). La capacità di eseguire esperimenti che forniscono dati in tempo reale su scala micron consente lo studio delle interazioni intra-specie e inter-specie come possono verificarsi in vivo. Ad esempio, SCFM2 è stato precedentemente utilizzato per studiare le dinamiche spaziali della comunicazione cellula-cellula in popolazioni aggregate attraverso una rete di sistemi utilizzati da Pa per regolare più geni che contribuiscono alla virulenza e alla patogenesi6.
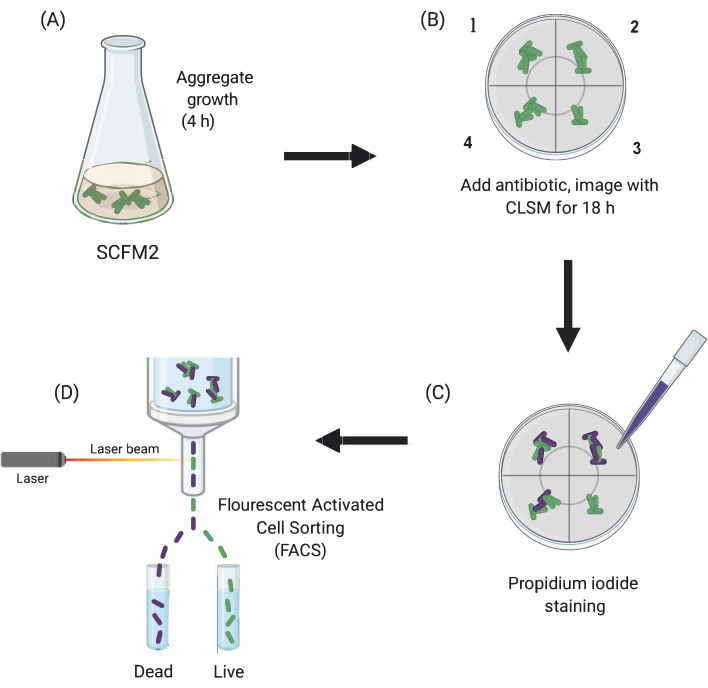
Figura 1: Rappresentazione grafica delle principali fasi sperimentali. (A) SCFM2 viene inoculato con cellule Pa e lasciato formare aggregati in un piatto di coltura con fondo di vetro. (B) Gli aggregati vengono trasferiti al microscopio confocale e viene aggiunto l’antibiotico. Sono raffigurate tre repliche tecniche (camere 1-3) e un pozzo di controllo (4) di SCFM2 inoculato senza trattamento antibiotico. Gli aggregati vengono ripresi utilizzando CLSM nel corso di 18 ore. (C) Dopo l’imaging iniziale di 18 ore, gli aggregati vengono trattati con ioduro di propidio per visualizzare le cellule morte e ripresi utilizzando CLSM (D) Gli aggregati con fenotipo desiderato vengono separati da SCFM2 utilizzando FACS. Abbreviazioni: SCFM2 = mezzo espettorato sintetico della fibrosi cistica; Pa = Pseudomonas aeruginosa; CLSM = microscopia a scansione laser confocale; FACS = selezione cellulare attivata dalla fluorescenza. Fare clic qui per visualizzare una versione più grande di questa figura.
Qui, viene dimostrata l’utilità di SCFM2 per studiare l’impatto del trattamento antibiotico sugli aggregati di Pa in tempo reale, seguita dall’uso di un approccio di ordinamento cellulare per isolare popolazioni di aggregati con fenotipi distinti per l’analisi a valle (Figura 1).
| Amino acids | |||
| Alanine | Acr s Organics s Organics |
56-41-7 | |
| Arginine HCl | MP | 1119-34-2 | |
| Asparagine | Acrs Organics |
56-84-8 | Prepared in 0.5 M NaOH |
| Cystine HCl | Alfa Aesar | L06328 | |
| Glutamic acid HCl | Acrs Organics |
138-15-8 | |
| Glycine | Acrs Organics |
56-40-6 | |
| Histidine HCl H2O | Alfa Aesar | A17627 | |
| Isoleucine | Acrs Organics |
73-32-5 | |
| Leucine | Alfa Aesar | A12311 | |
| Lysine HCl | Alfa Aesar | J62099 | |
| Methionine | Acrs Organics |
63-68-3 | |
| Ornithine HCl | Alfa Aesar | A12111 | |
| Phenylalanine | Acrs Organics |
63-91-2 | |
| Proline | Alfa Aesar | A10199 | |
| Serine | Alfa Aesar | A11179 | |
| Threonine | Acrs Organics |
72-19-5 | |
| Tryptophan | Acrs Organics |
73-22-3 | Prepared in 0.2 M NaOH |
| Tyrosine | Alfa Aesar | A11141 | Prepared in 1.0 M NaOH |
| Valine | Acrs Organics |
72-18-4 | |
| Antibiotic | |||
| Carbenicillin | Alfa Aesar | J6194903 | |
| Day-of Stocks | |||
| CaCl2 * 2H2O | Fisher Chemical | C79-500 | |
| Dextrose (D-glucose) | Fisher Chemical | 50-99-7 | |
| 1,2-dioleoyl-sn-glycero-3-phosphocholine (DOPC) | Fisher (Avanti Polar Lipids) | 4235-95-4 | shake 15-20 min at 37 °C to evaporate chloroform |
| FeSO4 * 7H2O | Acrs Organics |
7782-63-0 | this stock equals 1 mg/mL, MUST make fresh |
| L-lactic acid | Alfa Aesar | L13242 | pH stock to 7 with NaOH |
| MgCl2 * 6H2O | Acrs Organics |
7791-18-6 | |
| N-acetylglucosamine | TCI | A0092 | |
| Prepared solids | |||
| Porcine mucin | Sigma | M1778-100G | UV-sterilize |
| Salmon sperm DNA | Invitrogen | 15632-011 | |
| Stain | |||
| Propidium iodide | Alfa Aesar | J66764MC | |
| Salts | |||
| K2SO4 | Alfa Aesar | A13975 | |
| KCl | Alfa Aesar | J64189 | add solid directly to buffered base |
| KNO3 | Acrs Organics |
7757-79-1 | |
| MOPS | Alfa Aesar | A12914 | add solid directly to buffered base |
| NaCl | Fisher Chemical | S271-500 | add solid directly to buffered base |
| Na2HPO4 | RPI | S23100-500.0 | |
| NaH2PO4 | RPI | S23120-500.0 | |
| NH4Cl | Acrs Organics |
12125-02-9 | add solid directly to buffered base |
| Consumables | |||
| Conical tubes (15 mL) | Olympus plastics | 28-101 | |
| Conical tubes (50 mL) | Olympus plastics | 28-106 | |
| Culture tubes w/air flow cap | Olympus plastics | 21-129 | |
| 35 mm four chamber glass-bottom dish | CellVis | NC0600518 | |
| Luria Bertani (LB) broth | Genessee Scientific | 11-118 | |
| Phosphate-buffered saline (PBS) | Fisher Bioreagents | BP2944100 | |
| Pipet tips (p200) | Olympus plastics | 23-150RL | |
| Pipet tips (p1000) | Olympus plastics | 23-165RL | |
| Serological pipets (5 mL) | Olympus plastics | 12-102 | |
| Serological pipets (25 mL) | Olympus plastics | 12-106 | |
| Serological pipets (50 mL) | Olympus plastics | 12-107 | |
| Ultrapure water (RNAse/DNAse free); nanopure water | Genessee Scientific | 18-194 | Nanopure water used for preparation of solutions in Table 1 |
| Syringes (10 mL) | BD | 794412 | |
| Syringes (50 mL) | BD | 309653 | |
| 0.22 mm PES syringe filter | Olympus plastics | 25-244 | |
| PS cuvette semi-mico | Olympus plastics | 91-408 | |
| Software | |||
| Biorender | To prepare the figures | ||
| FacsDiva6.1.3 | Becton Dickinson, San Jose, CA | ||
| Imaris | Bitplane | version 9.6 | |
| Zen Black | |||
| Equipment | |||
| FacsAriallu | Becton Dickinson, San Jose, CA | ||
| LSM 880 confocal laser scanning microscope | Zeiss |
