מספר רב של אירועי תנועתיות בתוך תאים אאוקריוטים מתווכים על ידי תפקודם של חלבונים מוטוריים מולקולריים. מנועים אלה נעים לאורך חוטי השלד הציטוסקטליים, חוטי האקטין והמיקרוטובולים (MTs), וממירים את האנרגיה הכימית של הידרוליזה של ATP לכוחות קינטיים ומכניים הנדרשים להנעת תנועתיות ביולוגית בתוך התאים. S. cerevisiae Cin8 מבוסס MT הוא חלבון מוטורי דו-קוטבי, הומו-טרמרי קינזין-5, אשר מצליב ומחליק MTs צירים זה מזה1. Cin8 מבצע פונקציות חיוניות במהלך מיטוזה, בהרכבת ציר 2,3,4 והתארכות ציר במהלךאנפאזה 5,6,7. בעבר הוכח כי Cin8 הוא מנוע דו-כיווני, המחליף כיווניות בתנאי ניסוי שונים. לדוגמה, בתנאי חוזק יוני גבוה, מנועי Cin8 בודדים נעים לעבר הקצה התחתון של ה- MTs, בעוד שבאשכולות, במבחני גלישה MT מרובי מנועים, ובין MTs אנטי-פאראליים, מנועי Cin8 נעים בעיקר לעבר הקצוות החיוביים של MTs 8,9,10,11,12 . ממצאים אלה היו מאוד לא צפויים בגלל כמה סיבות. ראשית, Cin8 נושא את התחום המוטורי הקטליטי שלו בתחום האמינו-טרמינוס, ובעבר האמינו שמנועים כאלה מכוונים אך ורק ל-plus-end, בעוד ש-Cin8 הוכח כמינוס-קצה המכוון ברמת המולקולה הבודדת. שנית, מנועי קינסין נחשבו לחד-כיווניים, או מינוס-קצה או פלוס-סוף מכוון, בעוד ש-Cin8 הוכח כדו-כיווני, בהתאם לתנאי הניסוי. לבסוף, בגלל כיוון ה-MT בציר המיטוטי, התפקיד הקלאסי של מנועי קינסין-5 בהפרדת מוטות ציר במהלך מכלול הציר ואנאפאזה B יכול להיות מוסבר רק על ידי תנועתיות מכוונת פלוס-קצה שלהם על ה-MTs שהם מצליבים 1,13. בעקבות הדיווחים הראשונים על הדו-כיווניות של Cin8, כמה מנועי קינסין אחרים הודגמו כדו-כיווניים 14,15,16, מה שמצביע על כך שהתנועתיות הדו-כיוונית של מנועי קינסין עשויה להיות נפוצה יותר ממה שסברו קודם לכן.
בעבר דווח כי בתאים, Cin8 נע גם באופן דו-כיווני8, מה שתומך ברעיון שהתנועתיות הדו-כיוונית של חלק ממנועי קינסין-5 חשובה לתפקודים התוך-תאיים שלהם. בנוסף, מכיוון ששלושת מנועי הקינזין-5 שדווחו כדו-כיווניים הם מתאים פטרייתיים, הוצע לאחרונה תפקיד אפשרי לדו-כיווניות של מנועי קינסין-5 בתאים כאלה10. על פי מודל זה, במיטוזה סגורה של תאים פטרייתיים, שבה המעטפת הגרעינית אינה מתפרקת במהלך מיטוזה, מנועי קינסין-5 מספקים את הכוח הראשוני המפריד בין קטבי הציר לפני הרכבת הציר. כדי לבצע משימה זו, לפני הפרדת מוטות הציר, מנועי קינזין-5 מתמקמים ליד מוטות הציר, על ידי תנועתיות מכוונת מינוס-קצה שלהם על MTs גרעיניים בודדים. ברגע שהם נמצאים במיקום זה, מנועי kinesin-5 מקבצים, מחליפים כיווניות, לוכדים ומקשרים MTs צולבים מעמודי ציר שכנים. לאחר מכן, מנועי kinesin-5 מספקים את ההפרדה הראשונית של הקטבים על ידי תנועתיות מכוונת פלוס-קצה על ה-MTs שהם מצליבים. על פי דגם זה, הן תנועתיות מכוונת מינוס-קצה על MTs בודדים והן תנועתיות מכוונת פלוס-קצה על MTs מקושרים צולבים במהלך החלקה אנטי-פראלית נדרשים למנועי קינסין-5 פטרייתיים כדי לבצע את תפקידם בהרכבת ציר 1,13.
המטרה הכוללת של השיטה המתוארת היא להשיג קינסין-5 Cin8 פטרייתי בעל טוהר גבוה, ולבצע מבחני תנועתיות של מולקולה בודדת (איור 1) תוך ניתוח נפרד של התנועתיות של מולקולות בודדות ואשכולות של Cin8. ההפרדה בין מולקולות בודדות לאשכולות חשובה מכיוון שאחד הגורמים שהוכחו כמשפיעים על הכיווניות של Cin8 הוא הצטברותו באשכולות על MTs10,12. מבחני תנועתיות חלופיים, כגון מבחני החלקה על פני השטח של MT ומבחני הזזת MT אינם מספקים מידע לגבי פעילותם של חלבונים חד-מנועיים17,18. שיטות הבדיקה והניתוח החזקות של תנועתיות מולקולה בודדת המתוארות כאן יושמו בהצלחה כדי לאפיין היבטים שונים של מנועי קינסין-5, Cin8 ו- Kip1 10,11,12,14,19,20.
כאן מוצג פרוטוקול מפורט לביטוי יתר וטיהור של Cin8, פילמור של MTs ובדיקת תנועתיות של מולקולה אחת. יתר על כן, מתוארים גם הניתוחים להבחנה בין מולקולות בודדות לאשכולות של Cin8, ולקביעת מהירויות מנוע בודדות ואשכולות על ידי ניתוח תזוזה ממוצעת (MD) וניתוח תזוזה ריבועית ממוצעת (MSD). פרוטוקול זה נועד לסייע לחוקרים לדמיין את כל שלבי ההליכים ולסייע בפתרון בעיות מסוג זה של בדיקות.
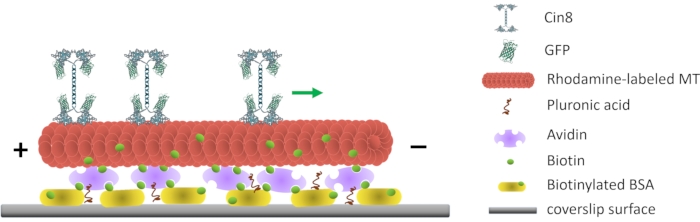
איור 1: ייצוג סכמטי של מבחן תנועתיות המולקולה הבודדת. MTs פלואורסצנטיים ביוטינילטיביים מחוברים לפני השטח של הזכוכית, מצופה באבידין המקיים אינטראקציה עם הביוטינילציה-BSA המחוברת לפני השטח. החץ הירוק מייצג את כיוון התנועה של מולקולות Cin8 בודדות בתנאי חוזק יוני גבוהים. +/- מייצגים את הקוטביות של MT. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
