A myriad of models are available to study the progression of cancer, each of them being unique and representing a subtype of this complex disease. Each model provides unique and valuable insights into cancer biology and has improved the means to mimic the actual disease condition. Established cell lines grown as a monolayer have provided valuable insights into vital processes in vitro, such as proliferation, invasiveness, migration, and apoptosis1. Though two-dimensional (2D) cell culture has been the traditional tool to investigate the response of mammalian cells to several environmental perturbations, extrapolation of these findings to predict tissue-level responses does not seem sufficiently convincing. The major limitation of the 2D cultures is that the microenvironment created differs largely from that of the breast tissue itself2. 2D culture lacks the interaction of the cells with the extracellular matrix, which is vital for the growth of any tissue. Also, tensile forces experienced by the cell in monolayer cultures hinder the polarity of these cells, thus altering cell signaling and behavior3,4,5. Three-dimensional (3D) culture systems have opened up a new avenue in the field of cancer research with their ability to mimic the in vivo conditions in vitro. Many crucial microenvironmental cues that are lost in 2D cell culture could be re-established using 3D cultures of laminin-rich extracellular matrix (lrECM)6.
Various studies have identified the importance of the tumor microenvironment in carcinogenesis7,8. Inflammation-associated factors are a major part of the microenvironment. Platelet Activating Factor (PAF) is a phospholipid mediator secreted by various immune cells that mediates multiple immune responses9,10. High levels of PAF are secreted by different breast cancer cell lines and are associated with enhanced proliferation11. Studies from our lab have shown that the prolonged presence of PAF in acinar cultures leads to the transformation of breast epithelial cells12. PAF activates the PAF receptor (PAFR), activating the PI3K/Akt signaling axis13. PAFR is also reported to be associated with EMT, invasion, and metastasis14.
The present protocol demonstrates a model system to study PAF-induced transformation, using 3D cultures of breast epithelial cells, as has been previously described by Chakravarty et al.12. The breast epithelial cells grown on the extracellular matrix (3D cultures) tend to form polarized growth-arrested spheroids. These are called acini and closely resemble the acini of breast tissue, the smallest functional unit of the mammary gland, in vivo15. These spheroids (Figure 1A,B) consist of a monolayer of closely packed polarized epithelial cells surrounding a hollow lumen and attached to the basement membrane (Figure 1C). This process of morphogenesis has been well described in literature16. When seeded on lrECM, the cells undergo division and differentiation to form a cluster of cells, which then polarize from Day 4 onwards. By Day 8, the acini consist of a group of polarized cells that are in direct contact with the extracellular matrix and a cluster of unpolarized cells enclosed within the outer polarized cells, with no contact to the matrix. These unpolarized cells are known to undergo apoptosis by Day 12 of culture, forming a hollow lumen. By Day 16, growth-arrested structures are formed16.
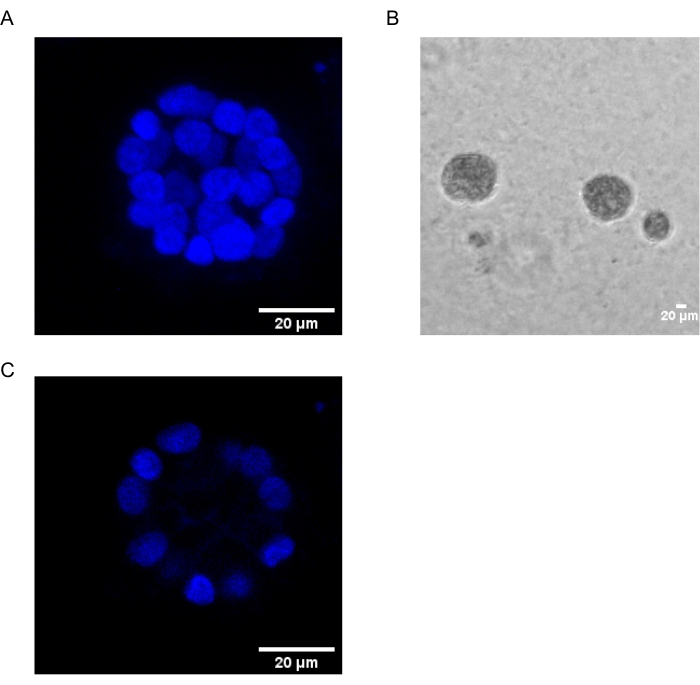
Figure 1: Nuclei of cells in acini stained with a nuclear stain. (A) 3D construction of the acini. (B) Phase Contrast image of MCF10A acini grown on Matrigel for 20 days. (C) The centermost section shows the presence of a hollow lumen. Scale bar = 20 µm. Please click here to view a larger version of this figure.
Unlike 2D cultures, acinar cultures aid in distinguishing normal and transformed cells through apparent morphology changes. Non-transformed breast epithelial cells form acini with a hollow lumen, mimicking the normal human breast acini. These spheroids, upon transformation, show a disrupted morphology characterized by a major loss of polarity (one of the hallmarks of cancer), absence of a lumen, or disruption of the hollow lumen (due to evasion of apoptosis) that may be induced due to deregulation of various genes17,18,19,20. These transformations can be studied using commonly used techniques such as immunofluorescence. Thus, the 3D cell culture model can function as a simple method to investigate the process of breast acinar morphogenesis and breast carcinogenesis. Establishing a 3D culture system to understand the effect of a phospholipid mediator, PAF, will assist in high throughput preclinical drug screening.
This work has adapted the 3D 'on top' culture protocol16,21 for studying transformation induced by PAF22. The phenotypic changes induced by exposure of the acini to the phospholipid mediator were studied using immunofluorescence. Various polarity and epithelial to mesenchymal transition (EMT) markers12,16 were used in the study. Table 1 mentions their normal localization and their expected phenotype upon transformation.
| Antibodies | Marks | Normal localization | Transformed phenotype |
| α6-Integrin | Basolateral | Basal with weak lateral stain | Strong lateral / Apical stain |
| β-Catenin | Cell-cell junction | Basolateral | Abnormal / nuclear or cytoplasmic localization |
| Vimentin | EMT | Absent / weak presence | Up-regulation |
Table 1: Markers used in the study. Different markers used with their localization in the presence and absence of PAF treatment.
This method can be best utilized to study/screen plausible drugs and target genes for various breast cancer subtypes. This can provide a drug response data closer to the in vivo scenario, aiding in faster and more reliable drug development. Also, this system can be used to study the molecular signaling associated with drug response and drug resistance.
