All animal experiments were conducted in accordance with the guidelines outlined by the National Research Council's Guide for the Care and Use of Laboratory Animals. Animal handling and euthanasia protocols were reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of the University of Illinois at Chicago.
1. Animal Models
- Wild-type Long-Evans rats
- Procure a 24 – 32 day old wild-type Long Evans Hooded rat of either sex raised with a standard 12 h day/night rhythm.
- Dark adapt the rat by placing it in a completely dark room for 1 h prior to beginning experiment.
- S334ter-3 rats
- Cross the transgenic albino homozygous S334ter-3 rat line (either sex), expressing two copies of the mutant rhodopsin gene, with a pigmented wild-type Long-Evans rat to produce pigmented heterozygous S334ter-3 rats that exhibit photoreceptor degeneration similar in progression to human retinitis pigmentosa29,30.
- Raise heterozygous offspring with standard 12 h day/night rhythm and use rats of either sex for experiments of the following ages corresponding to the following photoreceptor degeneration stages: Early stage degeneration: 14 – 20 days old; Middle stage degeneration: 21 – 27 days old; Late stage degeneration: 28 - 35 days old; Completely blind: >50 days old.
- Dark adapt the rat by placing it in a completely dark room for 1 h prior to beginning experiment.
2. Preparation of Ames' Medium Solution and Perfusion System
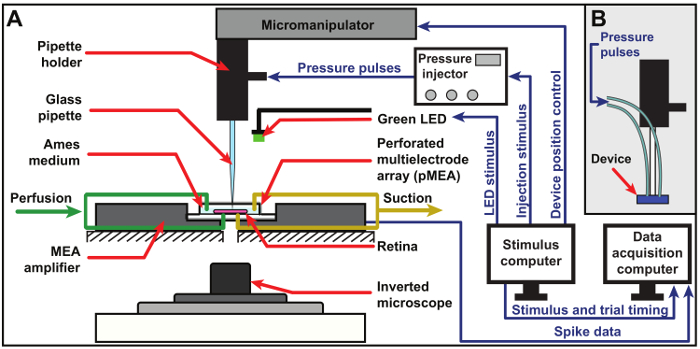
Figure 1: Schematic of experimental setup. Schematic of the experimental setup for chemical stimulation using a glass micropipette (A) and a custom multiport microfluidic device (B). The retina is placed on a pMEA and continuously perfused with fresh, oxygenated Ames medium solution from both the top and bottom through the pMEA perforations. Neural response signals picked up by the electrodes of the pMEA are fed through the MEA amplifier into a data acquisition computer. Visual and chemical stimulation are accomplished using a green LED and an 8-channel pressure injector, respectively, and both stimuli are triggered by a dedicated stimulus computer, which is also used to position the pipette via a precision 3-axis micromanipulator. An inverted microscope is used to observe the retina during an experiment. Please click here to view a larger version of this figure.

Figure 2: Experimental setup. (A) Photograph of the complete experimental setup showing the relative positions of all components. The MEA amplifier system is placed on top of an inverted microscope, which is used to visually inspect the retina and digitally image the device-retina interface by means of the attached high-speed camera during the experiment. Top and bottom perfusion are independently controlled using a solenoid-controlled perfusion system. Injection, position control, and impedance measurements are accomplished using a pressure injector, micromanipulator, and patch clamp amplifier (orange box; shown in more detail in B), respectively. The micropipette or device is inserted into a patch clamp headstage for impedance measurements and mounted on a gantry (indicated by green box; shown in more detail in C) to facilitate positioning using a micromanipulator. (B) A close-up of the measurement and control instruments used in the experiment: the 8-channel pressure injector, micromanipulator control system, patch clamp amplifier for impedance measurement, and the suction vessel for perfusion elimination. (C) A close-up of the injection system gantry showing a multiport device interfaced with the pipette holder, patch clamp headstage, and micromanipulator. Please click here to view a larger version of this figure.

Figure 3: Perfusion setup. (A) Photograph of the top of the MEA amplifier showing the location of the top perfusion and suction as well as the green LED used for visual light stimulation. (B) A close-up of the pMEA perfusion chamber illustrating the precise locations of the top perfusion and suction, the pMEA, and the reference electrode used for impedance measurement. (C) A photomicrograph of the bottom perfusion plate. Please click here to view a larger version of this figure.
NOTE: See Figure 1, Figure 2, and Figure 3
- Measure out 900 mL of de-ionized water at room temperature (~21 °C) and place in 1 L container.
- Perfuse the water with 100% CO2 using a bubbling mechanism.
- Add 8.8 g of powdered Ames' medium to water in 1 L container.
- Rinse Ames' medium container with a few mL of de-ionized water to remove all traces of powdered Ames' medium and add to 1 L container.
- Add 25.3 mL of sodium bicarbonate solution (7.5% w/v31) to 1 L container.
- Add additional water to bring the solution to a final volume of 1 L.
- Continue perfusing the water with CO2 for approximately 5 min.
- Stop CO2 perfusion and begin perfusing solution with a medical-grade gas mixture of 95% O2 and 5% CO2 for at least 30 min or until the pH stabilizes at 7.4.
NOTE: For the purposes of this protocol, the Ames medium is kept at room temperature (~21 °C) throughout the experiment to prevent CO2 or O2 from outgassing, which can occlude the perfusion lines with air bubbles. - Clean the bottom and top perfusion tubes by filling with 70% ethanol and then wash both lines 3 times with de-ionized water. Fill the bottom line with de-ionized water and the top line with air. Close both lines using a solenoid valve system.
- Attach the main perfusion tube to the luer connection of the 1 L Ames medium container.
- Open the top perfusion valve and leave it open until solution exits from the top perfusion outlet. Turn off the top perfusion valve.
- Open the bottom perfusion outlet and leave it on until all bubbles exit through the bottom perfusion outlet.
- Attach an empty suction vessel to the main suction line and turn on the suction source. Ensure that both the top and bottom suction inlets are open and working.
- Ensure that all computer displays are covered by red filter screens to avoid unintentional visual stimulation of the retina.
3. Wholemount Retinal Preparation
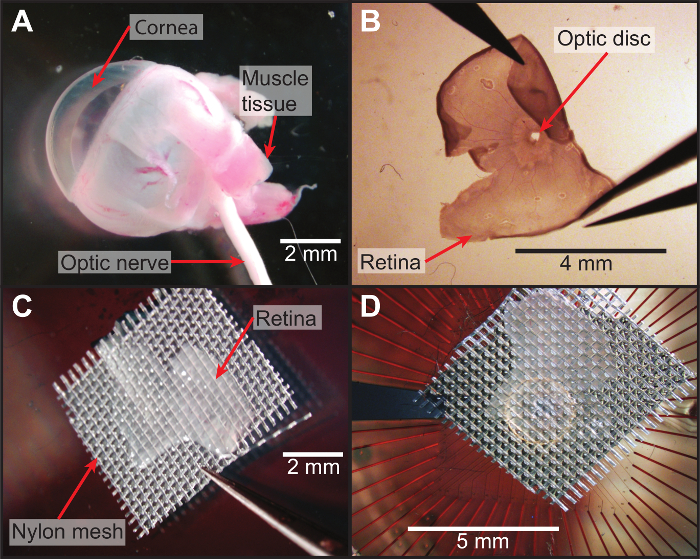
Figure 4: Dissection and wholemount preparation of retina. (A) Photograph of an intact eyecup taken from a photoreceptor-degenerated animal. (B) Photomicrograph of the retina with longitudinal cuts to flatten it out. (C) After flattening the retina, it is placed onto a mesh grid with the photoreceptor side contacting the mesh and flattened in air (outside of perfusion medium) to ensure there are no folds or curled edges. (D) The mesh and retina are quickly transferred to the pMEA with the ganglion cell side contacting the electrodes and immediately perfused with oxygenated Ames medium. Please click here to view a larger version of this figure.

Figure 5: Perforated multielectrode array. (A) Photograph of the perforated multielectrode array used in the protocol. The retina is placed on the electrode array (indicated by the red rectangle, which is shown in greater detail in B) within the pMEA chamber to allow continual perfusion with oxygenated Ames medium. (B) A photomicrograph of the electrode array itself illustrating the arrangement of perforations between electrodes. Please click here to view a larger version of this figure.
NOTE: See Figure 4 and Figure 5
- Using a handheld red LED flashlight to provide dim red illumination, euthanize animal via carbon dioxide asphyxiation followed by cervical dislocation or another chosen method, according to IACUC protocols.
- Enucleate both eyes using a jeweler's #5 forceps and place enucleated eyes in a 60 mm diameter petri dish with approximately 3 – 4 mL of fresh, oxygenated Ames medium solution.
- While observing the eye through a dissection stereomicroscope with top and bottom illuminators covered with red filter screen, make a small incision in the corneal face using a scalpel or a pair of sharp scissors.
- Cut from this small incision to edge of the cornea then extend the cut in a circumferential section around entire edge of cornea. Remove the now detached cornea along with the lens, translucent aqueous and vitreous humors.
- While gently holding the eyecup with one pair of forceps, carefully make two small incisions on opposite sides of the eyecup. Next, use two pairs of forceps to gently pull apart the eyecup at each of the incisions and separate the retina from the sclera. Slowly lift the entire retina from the sclera and eyecup. Cut the optic nerve, if it is still attached.
- Make longitudinal cuts in the retina to obtain half or quarter sections using scissors, and then gently spread one retinal section onto a nylon mesh (100 µm thread diameter with 350 µm opening) with the ganglion cells (concave side of retina) facing away from the mesh.
- Place the mesh and retina onto a perforated multielectrode array (pMEA) with the ganglion cells in contact with the pMEA surface. Then gently place a weighted slice grid on top of the mesh to hold the retina in place.
4. MEA and Data Acquisition Setup
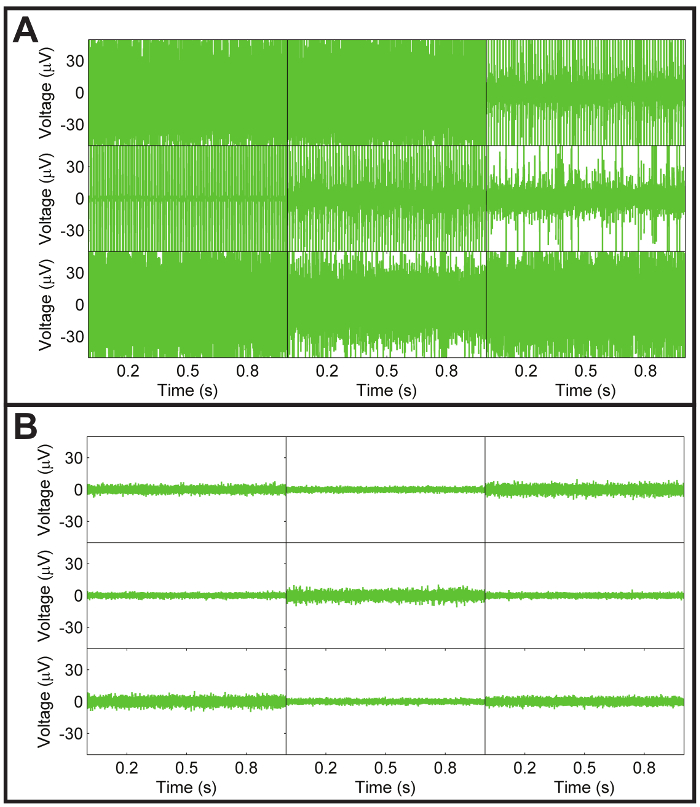
Figure 6: Noise levels of pMEA. (A) Representative recording of a subset of pMEA electrodes exhibiting high persistent noise. This noise is usually due to a lack of proper contact between the pMEA contact pads and the pins of the MEA amplifier as a result of normal wear of the thin perforated polyimide layer over the pMEA, especially at the contact pads. The other possible source of noise is typically solution leaked onto the pMEA amplifier contact pads. Persistent noise due to poor pin contact and/or leaked solution on the contact pads can usually be corrected by shifting the position of the pMEA within the amplifier to obtain better contact and cleaning and drying the pads, respectively. If noise cannot be eliminated by cleaning or shifting the pMEA position, the pMEA may need to be replaced entirely. (B) Representative recording of noise levels from a subset of pMEA electrodes from a clean pMEA with good contact between the pMEA and amplifier. Typically, the average noise level is within ±16 µV. Please click here to view a larger version of this figure.
NOTE: See Figure 6
- Under dim red illumination, place pMEA in MEA amplifier and close amplifier latches.
- If reference marks are not already present on the pMEA chamber ring, etch two 'X'-shaped marks spaced approximately 5 mm apart in an area easily visible from above with the boom-stand mounted microscope.
- Position the top perfusion outlet inside the pMEA chamber and turn on the top perfusion valve to achieve a perfusion rate of approximately 3 mL per minute. Position the top suction inlet at the desired perfusate level (~5 mm deep) and ensure that it is working.
- Open the data acquisition software on the data acquisition computer and click the 'play' button to start receiving data. Ensure that all pMEA channels are noise-free and recording neural signals. If not, reposition the pMEA within the amplifier to obtain better contact between the amplifier pins and the pMEA contacts (see Figure 5).
- Ensure that the bottom perfusion line is clear of any air bubbles and, if it is bubble-free, turn on the bottom perfusion valve to ensure the bottom of the retina is supplied with oxygen and nutrients.
- Turn on the high-speed camera attached to the inverted optical microscope (10X magnification with N.A. of 0.45) and open imaging software. Ensure that the inverted microscope illuminator is covered by a red filter sheet to emit only red light and then set the illuminator to a low light level to avoid photobleaching the retina.
- By looking at a live digital image of the inverted microscope field of view on a monitor, observe the bottom surface of the pMEA for evidence that solution is flowing through the bottom perfusion plate.
- Once bottom perfusion is confirmed to be flowing, slowly ramp up the bottom suction by manually turning the vacuum pressure knob on the vacuum waste kit while observing the retina through the inverted microscope. Cease increasing the suction once an observable suction force acts on the retina. Be careful to avoid too much or too little suction.
- After ensuring the bottom suction holds the retina in place, take the weighted slice grid off the retina using forceps and gently remove the nylon mesh by peeling one corner carefully from the retina. It should separate easily leaving the retina firmly attached to the bottom of the pMEA with the subretinal surface exposed on top. Keep perfusion running for approximately 30 min to allow retina to stabilize from surgical trauma.
5. Glutamate Stimulation Preparation

Figure 7: Glass micropipette and multiport microfluidic device. (A) A pipette holder before inserting a micropipette or device. The silver/silver chloride electrode (50 µm diameter) is electrically coupled to the adapter on the end to interface with the patch clamp headstage. (B) A photograph of the glass micropipette interfaced with the pipette holder showing the location of the pressure port used to initiate pneumatic injections. (C) A custom multiport microfluidic device (1 cm 1 cm 0.134 cm) attached to a custom 3D-printed fixture and interfaced with the pipette holder through a stainless-steel tube. The device, which was fabricated in two layers, has eight microports (diameter 14 µm) in the bottom layer (340 µm thick) and eight on-chip reservoirs (diameter 1.6 mm) for storing glutamate in the top layer (1 mm thick). Each of the eight microports in the bottom layer of the device is independently connected to an on-chip reservoir in the top layer via an in-plane microchannel, and each on-chip reservoir in turn is connected to a pressure port of the 8-channel pressure injector via a flexible tube to allow independent actuation of the microports for patterned multisite injections. (D) A close-up of the multiport device held by tweezers before attaching the tubing interface fixture showing the arrangement of the eight independently-addressable on-chip reservoirs and tubing inlets. (E) A photomicrograph of the bottom surface of the device showing the eight 14 µm-diameter microports arranged in a 3 x 3 configuration with 200 µm spacing to align with the electrodes of the pMEA. The eight outside microports are utilized for multisite injections, while the central port was used strictly for alignment during fabrication of the device. Please click here to view a larger version of this figure.
NOTE: See Figure 7.
- Prepare glutamate solution by mixing stock glutamate solutionwith oxygenated Ames medium solution to obtain a 0.5 mL sample at a working concentration of 1 mM glutamate.
- Carefully insert a pre-pulled 10 µm-diameter micropipette or the stainless steel rod connected to the multiport microfluidic device into a standard pipette holder containing a 50 µm-diameter silver/silver chloride wire electrode.
NOTE: If impedance detection is not available, the silver/silver chloride wire electrode may be omitted. - Interface the pipette holder with the patch-clamp headstage and connect the pressure port luer connection of the pipette holder to channel 1 of the pressure injection system, if utilizing a glass micropipette, or connect the pressure port luer connections of each of the 8 injection ports with channels 1 – 8 of the pressure injection system, if using the multiport device.
- Manually turn on the pressure injection system and turn on channel 1 (or channels 1 – 8, as applicable). Ensure that the system is vented to atmosphere and set the injection pressure to 0.1 psi.
- Turn on micromanipulator and calibrate it by pressing the 'Calibrate' button on the manipulator controller. Position the micromanipulator so that the micropipette tip (or the bottom of the device, as applicable) is approximately 30 mm above the MEA amplifier.
- Fill a small petri dish with glutamate solution (1 mM glutamate in standard Ames medium) and place it underneath the micromanipulator. Lower micropipette tip (or the device) into the solution and fill by pressing the 'Fill' button on the pressure injection system (suction pressure of -13 in H2O) until there is approximately 20 mm of solution visible in the glass micropipette or the multiport device tubing.
- If using the multiport device, turn channel 1 of the pressure injector off and repeat the protocol for channels 2 – 8. Lift the micropipette tip or device out of solution, remove the petri dish, and position the micromanipulator above the pMEA chamber.
- Using a boom-stand-mounted stereomicroscope, align the micropipette tip or the corners of the device with the reference marks etched into the pMEA chamber ring. Store the manipulator positions into the control software using the 'Store Reference A' and 'Store Reference B' buttons (or simply note the manipulator coordinates manually) to map the coordinate system of the manipulator with the pMEA electrodes.
- Using the manipulator control software, select a target pMEA electrode with robust spontaneous activity and click the 'Move to Channel' button to align the glass micropipettewith the target electrode. If using the multiport device, align the device microports with target pMEA electrodes with robust spontaneous activity using the same process.
6. Interface with Retina
- If impedance measurement is available, turn on patch clamp amplifier and initiate the impedance visualization software by clicking the 'Start' button to visualize the impedance of the silver/silver chloride electrode inside the pipette holder. While observing the real-time impedance signals, slowly lower the micropipette or device until it contacts the retinal surface as indicated by a rapid increase in the impedance signal (see Figure 8). Save or make note of the position of the retinal surface.
- If impedance measurement is unavailable, detect contact with the retinal surface through visual observation, though this will be less precise. Lower the pipette or device until it visibly contacts the top surface of the Ames medium solution in the MEA chamber.
- Then, while observing the top of the retina with an inverted microscope, slowly lower the pipette or device until the top surface of the retina is visibly distorted, which indicates that contact has been made with the retinal surface. Save or make note of the position of the retinal surface.
- For subsurface stimulation, lower the pipette a further 40 µm (for S334ter-3 retinas) or 70 µm (for wild-type retinas).
- Perform a few short duration (10 – 30 ms) injections using the pressure injection (0.1 psi) system to determine if the cells near the micropipette tip or device microports are receptive to glutamate stimulation by observing the neural signals with data acquisition software.
NOTE: Successful injections will elicit a clearly visible spike rate burst or spike inhibition (see Figure 9). If no response is observed, reposition the micropipette or device at a different electrode.
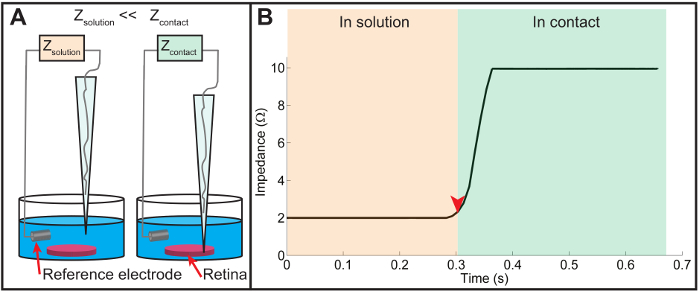
Figure 8: Impedance measurement. (A) A schematic of the impedance measurement technique. Using the patch clamp amplifier, the impedance of the micropipette is continuously monitored as it is slowly lowered towards the retinal surface. When the micropipette is above the retina, the relatively high ionic conductivity of Ames medium results in a low impedance reading. As the micropipette makes contact with the retinal surface, the ionic conductivity through the silver/silver chloride wire is reduced, causing a rapid increase in measured impedance. (B) A plot displaying the impedance change recorded just before and after the contact of the pipette tip with the retinal surface. The measured impedance is relatively low when the micropipette tip is in solution just prior to contact (indicated by the orange region on the left). Once contact is made (indicated by the red arrowhead and the green region on right), the impedance rapidly increases due to reduced ionic conductivity upon contact with the retinal tissue. In practice, the retinal surface is registered as the height corresponding to the onset of the steep rise of the measured impedance (the location of the red arrowhead). Please click here to view a larger version of this figure.
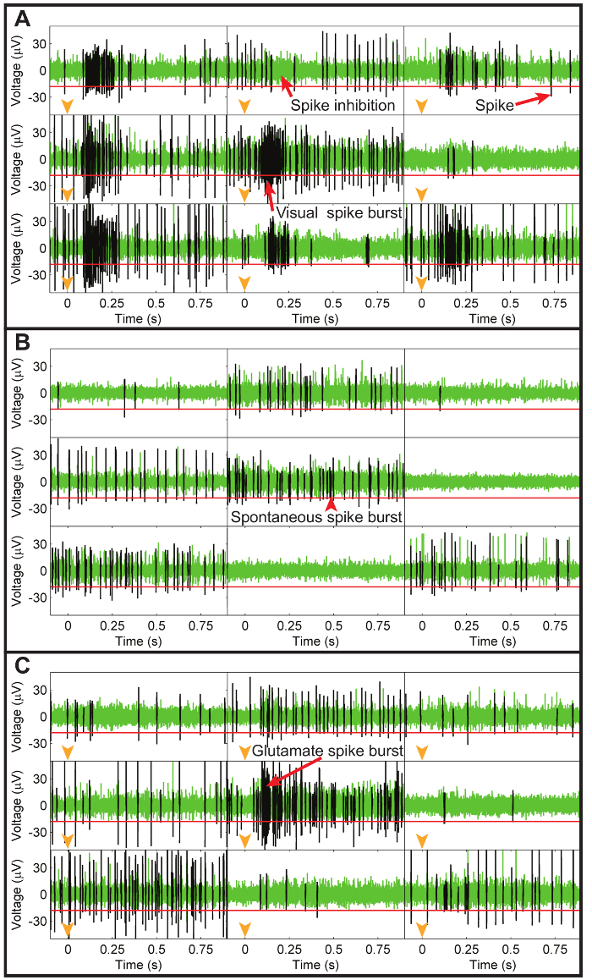
Figure 9: Electrode recordings of neural activity during visual, spontaneous, and glutamate injection recordings. (A) Representative recordings from nine pMEA electrodes showing the high-pass filtered electrode data during visual light stimulation with a green LED where each rectangle shows the neural data from a unique electrode. Each electrode recording illustrates data collected in the first second after turning on the green LED (timing shown with orange arrowheads in each plot) with a common voltage scale shown in the left y-axes. Spikes were identified using a threshold voltage of -18 µV (horizontal red line in each electrode plot) and are represented by the black traces over the green electrode data. Visual stimulation caused a burst of spikes (excitation) in all electrodes except the top center one, which possessed an inhibitory response to light. (B) A similar plot for the same electrodes showing spontaneous neural activity without visual or injection stimulation. Although smaller bursts were present, the patterns of spikes were very different from those recorded in response to visual stimulation. (C) Representative recordings from the same subset of electrodes recorded immediately after a glutamate injection at the central electrode (timing indicated by orange arrowheads in each plot). The injected glutamate elicited a burst of spikes in the central electrode that was very similar to the visually-evoked spike bursts. All other electrodes were unaffected by the glutamate injection, which demonstrates the fine spatial resolution of the chemical stimulation technique. Please click here to view a larger version of this figure.
7. Initiate Retinal Recording and Stimulus Program
- Orient the green LED toward the top surface of the retina. Begin recording using the data acquisition software on the dedicated recording computer by typing the filename and clicking the "record" button.
- Once recording has started, open the stimulus control program and load the default stimulus file by clicking the "Read Stimulus File" button. Next, click on the "Run Stimulus File" button to initiate the default stimulus file consisting of the stimuli and data acquisition protocol described in the note below.
NOTE: (i) 30 trials of 2 s ON and 2 s OFF full field flash (5 lm/m2 intensity) using the green LED. (ii) 120 s of data without using the green LED to record the spontaneous activity of the retina over a similar timescale. (iii) 1 or more sets of glutamate injections consisting of 30 trials of glutamate injections at 0.1 psi with 10 – 30 ms injection times (approximately 100 – 300 pL per injection) and 3 s interpulse durations. In the case of multi-site injections, select 2 or more ports to inject simultaneously. (iv) 90 s of spontaneous activity. - Once the stimulus file has been completed, stop the recording (by pressing the "Stop" button) to save the file for future spike sorting and data analysis (see Figure 10 for example).
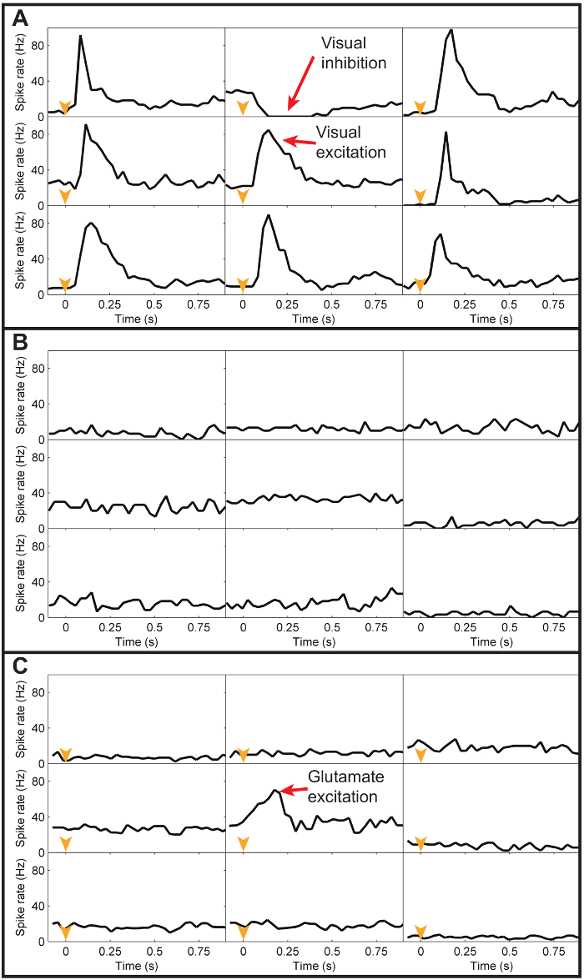
Figure 10: Peristimulus histograms of visual, spontaneous, and glutamate responses. (A) Representative peristimulus histograms (30 ms binwidth) of the spike data from the subset of electrodes in Figure 9A, averaged across 20 trials of visual light stimulation. A common spike rate scale was applied to all electrodes and is shown on the left y-axes. The black line in each electrode plot represents the average spike rate for all spikes recorded during the first second after turning the green LED on. As can be seen, visual stimulation caused a transient excitatory spike rate response at all electrodes except the top center electrode, which had a transient inhibitory response to light. (B) The average spontaneous spike rate responses recorded at the same electrodes without any visual or chemical stimulation. Without stimulation, the spike rates for each electrode are relatively constant. (C) The average spike rate responses recorded at the same electrodes in response to a glutamate injection at a location above the central electrode. The only transient response evident is the excitatory response at the electrode directly under the injection site. Please click here to view a larger version of this figure.
