In Figure 1, biofilm formation is induced in most of the six enteric pathogens tested following growth in media containing bile salts. A significant increase in adherent bacteria after bile salts exposure is observed in nearly all strains tested. The exception is enteroaggregative E. coli (EAEC); however, note the induced observation of the Δaaf mutant4. The results indicate that additional adherence mechanisms are induced by bile salts exposure in EAEC in the absence of aggregative adherence fimbriae I (AAF/I)21. To draw conclusions from this data set, plot the OD540 values for each strain and media type tested. Comparing the values from each strain in the media control (-BS) relative to media containing bile salts will determine if bile salts significantly induce biofilm formation. Comparisons across bacterial strains and/or mutants can also be performed for further characterization analyses.
The ConA-FITC assessment of EPS is presented in Figure 2. Figure 2A represents the semi-quantitative assessment of EPS production in which ConA-FITC is used to stain the biofilms. The ConA binds to the polysaccharides in the EPS matrix and the amount retained is detected by a fluorescent plate reader. To present the data, plot the average fluorescence units for each condition and compare the biofilm-inducing conditions to the negative control. Additionally, comparisons across bacterial strains or mutants can be performed. It is critical to analyze a media-only, negative control to determine the amount of background ConA-FITC fluorescence that occurs for the plate. The TSB condition was not significantly different from the media control, while the fluorescence readings significantly increased following bile salts exposure. The data confirm that EPS production is indicative of biofilm formation and does not occur in the absence of bile salts for S. flexneri. The images of bile salt-induced biofilms in S. flexneri is represented in Figure 2B. ConA-FITC is used to stain polysaccharides in the EPS matrix while a DAPI counterstain is used to visualize the bacteria by staining the DNA. For bacteria not exposed to bile salts (negative control), only DAPI is detected. Notably, the density of bacteria is much lower than in the bile salts condition. A clear background on the FITC channel is obtained to indicate the lack of EPS production. As expected, the data demonstrate that EPS production is correlated with biofilm formation1,2,3 following bile salts exposure. The -ConA staining control in the bile salt-treated samples is provided to demonstrate the specificity of the FITC fluorescence. For these images, 1x PBS was used in place of ConA-FITC and the DAPI counterstain was maintained. Imaging can be performed with a confocal microscope to assess the thickness of biofilms with the EPS-positive samples4.
Bacterial dispersion from biofilms in Figure 3 is detected by incubating established biofilms in PBS or supplemented PBS. Physiologically, bile is reabsorbed into circulation in the small intestine and only 5% of bile enters the colon5. As Shigella invades the colonic epithelium, we hypothesize that reabsorption of bile is an important signal for biofilm dispersion. In this figure, dispersion occurs in biofilms exposed to PBS or PBS + glucose; however, dispersion is inhibited in PBS + bile salts regardless of the presence of glucose. The data demonstrate that the removal of bile salts is necessary and sufficient to induce dispersion of S. flexneri from bile salt-induced biofilms4. To plot the data, either the recovered CFU of the bacteria or the relative percent to the control (as shown here) can be plotted.
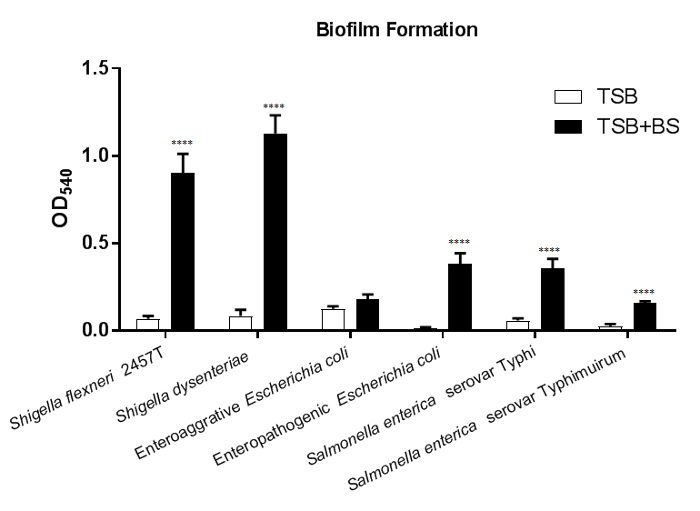
Figure 1: Bile salts induce biofilm formation in enteric pathogens. 18 h biofilms were grown on tissue-culture treated 96-well plates in either TSB or TSB + 0.4% bile salts media and stained with 0.5% crystal violet. Quantitative values from ethanol solubilization of the crystal violet stain were determined by spectrophotometry at OD540. Bile salts exposure significantly increased biofilm formation in Shigella flexneri, S. dysenteriae, enteropathogenic Escherichia coli, Salmonella enterica serovar Typhi, and S. enterica serovar Typhimurium. Statistical significance was determined by the student's T-test for each strain comparing the bile salts treatment to the media control. The standard error of the mean (SEM) is represented by the error bars. All p-values are <0.0001. Data shown are from one (n = 3) representative experiment, with a full set of data provided in a previous publication4. Please click here to view a larger version of this figure.
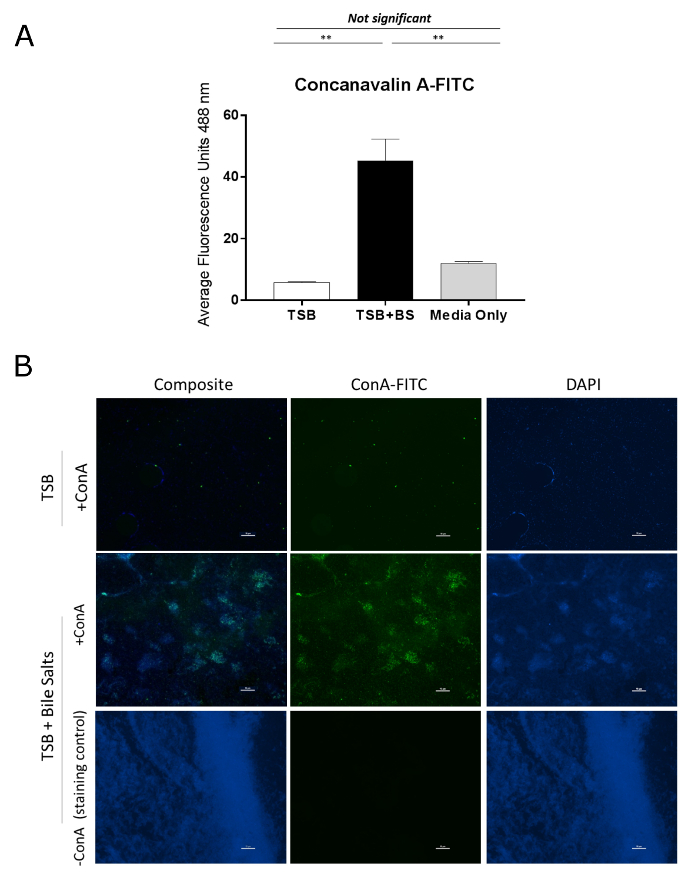
Figure 2: Bile salt-induced biofilm formation is characterized by production of a EPS matrix that is detected with FITC-labeled Concanavalin A. (A) Sterile black, 96-well plate were seeded with S. flexneri at a 1:50 dilution into TSB or TSB + bile salts and grown for 18 h. Biofilms were fixed, gently washed, and stained with ConA-FITC. The amount of retained Concanavalin-A FITC was determined by a fluorescent plate reader. The values plotted demonstrate EPS quantification following growth of S. flexneri grown in TSB or TSB-containing bile salts. Significance was determined by one-way ANOVA and p-values are <0.01. The SEM is represented by the error bars. Data shown are from one (n = 3) representative experiment, with a full set of data provided in a previous publication4. (B) Microscopy images from a bile salt-induced Shigella biofilm. Sterile coverslips were seeded with S. flexneri at a 1:50 dilution in TSB or TSB + bile salts and grown for 18 h. The coverslips were subsequently fixed and stained with ConA-FITC and/or counterstained with DAPI. ConA-FITC was only detected in the bile salts condition due to the presence of the EPS matrix. A coverslip stained only with DAPI in the TSB + bile salts treatment was used to demonstrate minimal background fluorescence for the FITC setting on the confocal microscope. Images for each wavelength channel and a composite overlay are provided for all treatment conditions. Scale bars indicate 10 µm. Data shown are from one (n = 3) representative experiment, with a full set of data provided in a previous publication4. Please click here to view a larger version of this figure.
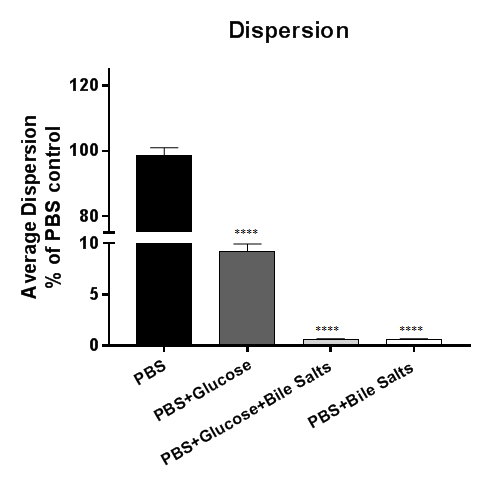
Figure 3: Analysis of bacterial dispersion from bile salt-induced biofilms. 18 h bile salt-induced Shigella flexneri biofilms were exposed to PBS or PBS supplemented with glucose, bile salts, or a combination of glucose and bile salts. The supernatant was subsequently serially diluted and plated on agar plates to enumerate the colony forming units (CFUs) that dispersed the biofilm. The number of bacteria recovered was plotted relative to the PBS control. Bacteria dispersed from the biofilm in the PBS or PBS + glucose conditions, but the presence of bile salts was sufficient to inhibit bacterial dispersion from the biofilm. Data shown are from one (n = 3) representative experiment, with a full set of data provided in a previous publication4. For reference, 100-fold more bacteria were routinely recovered from the PBS control treatment (approximately 7 x 107 CFU) compared to the PBS + bile salts treatment (approximately 7 x 105 CFU). Significance was determined by one-way ANOVA and p-values are <0.0001. The SEM is represented by the error bars.
