HAEC were isolated from each of the three anatomical regions of the amniotic membrane and individually cultured in vitro. After 48 h of culture, cells with an epithelial phenotype adhered to the surface of the plate, although the media also contained cell debris and floating cells, which were removed once the medium was changed (Figure 3).
During the processing of primary culture (passage zero, P0), some complications could arise that can interfere with the experimental data analysis (Figure 4): it is advisable to discard the cultures and process another membrane upon identifying the presence of bacteria due to contamination of the reagents or during the isolation process (Figure 4A); excessive erythrocytes due to insufficient washing of the membranes (Figure 4B); deficient or no adhesion of the cells to the plates (Figure 4C); or cells with fibroblast morphology (Figure 4D), suggesting that human amniotic mesenchymal cells (HAMC) were isolated instead of HAEC.
HAEC morphology depends on the origin of these cells: cells from the reflected zone have a cuboidal morphology and grow in a cobbled monolayer, unlike cells from the placental and umbilical regions, which are flatter and squamous (Figure 5). These data support that the epithelial layer from the amnion is not uniform throughout the membrane. During the passages, the size of the cells from all regions increases, but they maintain their epithelial nature and do not acquire a fibroblast morphology. Indeed, immunofluorescence against E-cadherin showed that the primary cultures (P0) and subcultures (P1-P2) maintained their epithelial phenotype (Figure 6), suggesting that there is no contamination of another cell type, such as mesenchymal stromal cells, or epithelial-mesenchymal transition. In addition, these cells show no evidence of cell death according to the TUNEL assay (Supplementary Figure 1) but are positive for the KI-67 proliferation marker (Figure 6), although our previous results found no significant differences for this marker between each passage5.
The number of cells obtained varies according to the region: 61.6 x 106 and 71.8 x 106 cells from the reflected and placental regions, respectively, and less than 1 x 106 per sample from the umbilical region (Table 1). It has been reported with this protocol that placental and umbilical region are very similar in their expression profiles, as opposed to the reflected and placental regions, especially in genes that participate in ECM receptor interaction, focal adhesion, and the PI3K-Akt signaling pathway through RNA-seq6. In agreement, a previous study reported the differential expression of mitogen-activated protein kinase and transforming growth factor beta pathways, as well as proinflammatory cytokines between both regions17.
Although the evidence showed that the subpopulations from the amnion differ in their morphology and physiological properties, we previously demonstrated that the expression and presence of the core of the pluripotency factors does not change in HAEC derived from placental and reflected regions6. In this context, in addition to the relatively limited number of cells from the umbilical region, subsequent studies should focus on isolated HAEC from the placental and reflected amnion independently, considering the physiological implications, because the different subpopulations would not respond equally to specific events, such as prolonged pregnancy or inflammatory processes during labor, although there are no specific positive cells to the main pluripotency markers (Figure 7).
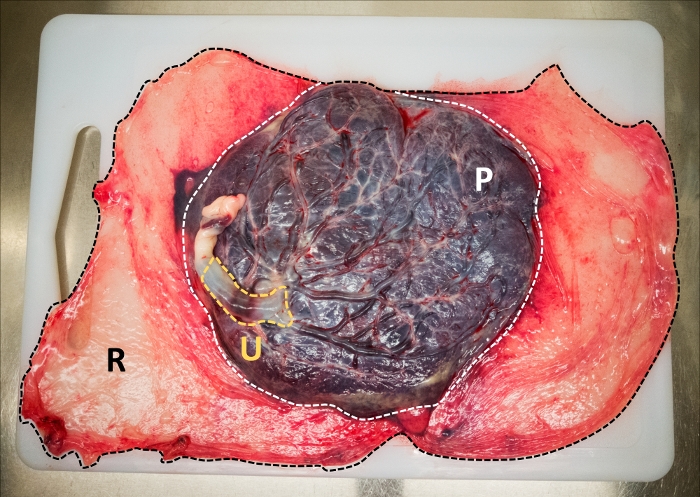
Figure 1: Anatomical regions from the amniotic membrane at term. Umbilical amniotic membrane (yellow), placental amniotic membrane (white), and reflected amniotic membrane (black). Please click here to view a larger version of this figure.
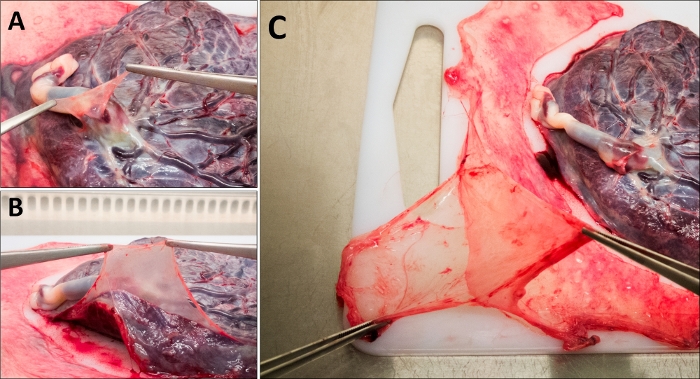
Figure 2: Mechanical dissection of the amniotic membrane. (A) Umbilical region, (B) placental region, and (C) reflected region. Please click here to view a larger version of this figure.
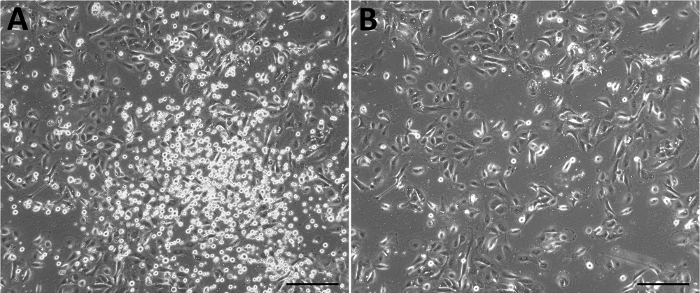
Figure 3: Representative primary culture of HAEC derived from the amniotic membrane. Culture 48 h after isolation without (A) and with (B) a change of media. Scale bars = 50 µm. Please click here to view a larger version of this figure.
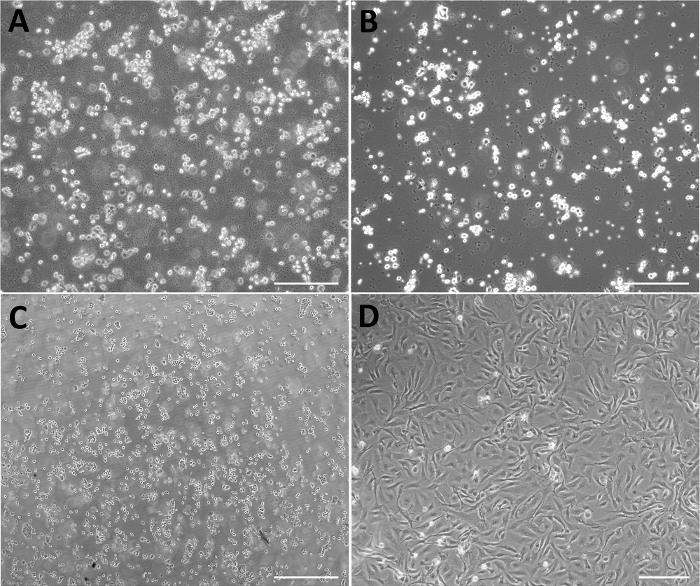
Figure 4: Representative micrographs of negative results of primary culture of HAEC. (A) Excess of erythrocytes. (B) Bacterial contamination. (C) HAEC not adhered after 48 h of isolation. (D) Primary culture composed of cells with fibroblast morphology. Scale bars = 200 µm. Please click here to view a larger version of this figure.

Figure 5: Morphology of HAEC from different anatomical regions in vitro. Representative micrographs of confluent HAEC from reflected (upper panel), placental (middle panel), and umbilical (lower panel) regions cultured though P0−P2. Scale bars 200 = µm. Please click here to view a larger version of this figure.
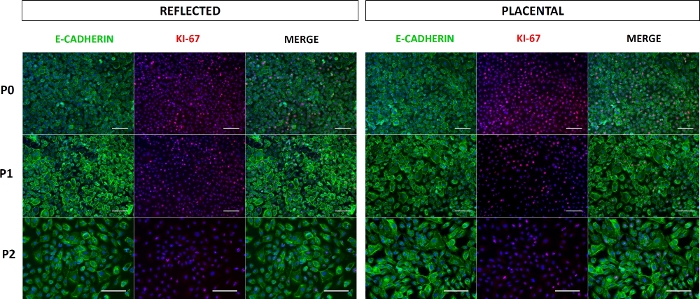
Figure 6: Expression of E-cadherin and KI-67 in HAEC. Representative epifluorescence microscopy images of E-cadherin+ and KI-67+ HAEC from reflected (left panel) and placental (right panel) amnion cultured through P0−P2. Scale bars = 100 µm. Please click here to view a larger version of this figure.
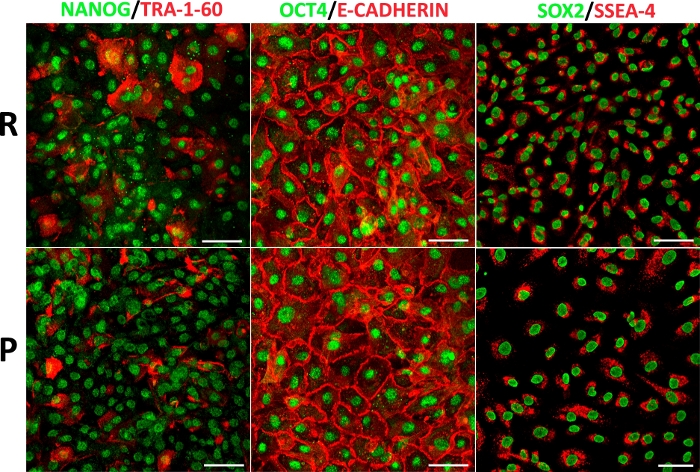
Figure 7: HAEC from different anatomical regions display a pluripotency marker panel. Representative confocal microscopy images of double immunofluorescence amnion for NANOG with TRA-1-60, OCT4 with E-cadherin, and SOX2 with SSEA-4 in HAEC (P1) from reflected (upper panel) and placental (lower panel) regions. Scale bars = 100 µm. Please click here to view a larger version of this figure.
| # MEMBRANE | REFLECTED | PLACENTAL | UMBILICAL | COMPLETE MEMBRANE |
| 1 | 75 X 106 | 130 X 106 | 0.2 X 106 | 205.2 X 106 |
| 2 | 38.5 X 106 | 52 X 106 | 0.53 X 106 | 91.03 X 106 |
| 3 | 59 X 106 | 53 X 106 | 0.2 X 106 | 112.2 X 106 |
| 4 | 42 X 106 | 27 X 106 | 0.36 X 106 | 69.36 X 106 |
| 5 | 44.8 X 106 | 22.3 X 106 | NP | 76.1 X 106 |
| 6 | 100 X 106 | 140 X 106 | 1 X 106 | 241 X 106 |
| 7 | 72 X 106 | 78 X 106 | NP | 150 X 106 |
| AVERAGE | 61.6 X 106 | 71.8 X 106 | 0.45 X 106 | 133.8 X 106 |
Table 1: Number of HAEC isolated per region from amniotic membranes. (NP = not processed).
Supplementary Figure 1: TUNEL staining in HAEC (passage 2) from reflected region and in positive control cells (HL-60 cells treated with camptothecin). Scale bars = 50 µm. Please click here to download this figure.
