Large-scale functional screening of Ube3a missense variants identifies a broad landscape of loss- and gain-of-function mutations
Previous work with Ube3a mutants suggested that the Wnt response can serve as a reporter of cellular UBE3A protein activity. These observations were expanded, and additional validation experiments were performed to investigate whether the BAR assay is suitable to report a range of UBE3A activities in the cell. First, HEK293T cells were transfected with varying amounts of plasmid DNA encoding human WT Ube3a. This experiment demonstrated that the BAR response changes linearly with the quantity of Ube3a transfected into cells (Figure 2A). Secondly, the BAR assay was tested using plasmid DNA encoding disease-linked variants previously characterized in the literature22. Among these were benign variants (R39H and A178T), one autism-linked gain-of-function variant (T458A), and a collection of confirmed Angelman syndrome-linked variants that cause a loss of UBE3A function through various mechanisms15,22. The BAR assay correctly identified each variant (Figure 2B), including all loss-of-function variants, demonstrating the accuracy and generality of this method18.
The BAR assay was used to screen 152 missense variants, most of which were identified from the ClinVar database. Similar to genetic evidence regarding copy number variations in Ube3a-dependent disorders, the functional impact of missense variants was broad and included benign variants as well as both loss-of-function and gain-of-function variants (Figure 3)18. Of particular interest, the gain-of-function variants identified using this method were all novel, and subsequent validation strongly suggested that they define a sub-class of Ube3a-dependent disorders with phenotypes that differ from classic Angelman syndrome18.
The effect of missense variants is often unpredictable, underscoring the need for functional assessment
An intriguing observation from the functional screening of Ube3a variants is that changes at some amino acid positions produce drastically different effects, thereby underscoring the importance of empirical variant assessment18. For example, three individuals were identified with variants at the glutamine 588 (Q588) position in UBE3A that included a gain-of-function glutamate substitution (Q588E), a loss-of-function arginine substitution (Q588R), and another loss-of-function proline substitution (Q588P; Figure 4A). A comprehensive mutation approach was used to mutate the Q588 position to every possible amino acid. When the activity of these variants was measured using the BAR assay, it showed that any negatively charged amino acid at the 588 position produces a hyperactive enzyme (Figure 4B). This insight provided important functional clues that allowed the discovery of a novel site within the HECT domain of UBE3A that facilitates ubiquitin chain elongation18.
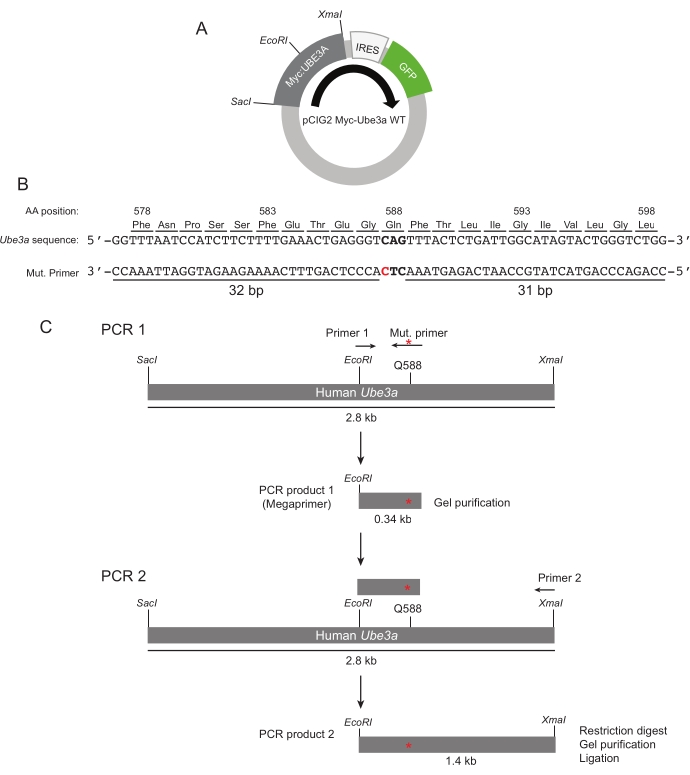
Figure 1: PCR-dependent mutagenesis of Ube3a. (A) Representation of the Ube3a expression vector indicating relevant restriction sites. (B) The DNA sequence encoding UBE3A is shown at the top. The Q588 codon is indicated by bold lettering. The Q588E mutagenesis primer (Mut. primer) is shown aligned to the UBE3A sequence. The red lettering indicates the mutagenized site. (C) A schematic of the PCR to generate the megaprimer and the DNA fragment used to subclone the Q588E fragment is shown. The position of the Q588E mutation is indicated by the red asterisk. Please click here to view a larger version of this figure.
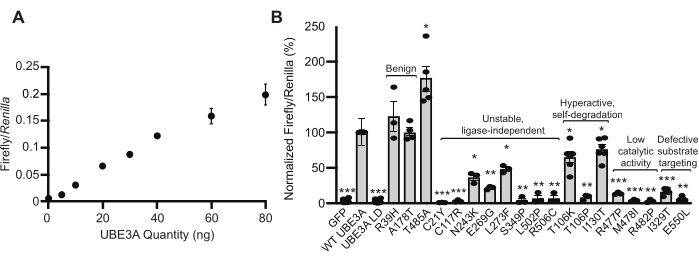
Figure 2: Validation of the BAR assay as a reporter of UBE3A ubiquitin ligase activity. (A) HEK293T cells transfected with increasing amounts of plasmid encoding WT UBE3A show a linear increase in the BAR response. Mean values are shown for Firefly/Renilla luciferase ratios ± standard error (SE). N= 3 independent experiments. (B) Previously characterized Ube3a variants were tested in the BAR assay. Responses were normalized to WT UBE3A, and the values are shown as the mean ± SE. The number of experiments (n) and p-values calculated using a one-sample t-test (two-tailed) with Benjamini-Hochberg multiple comparisons correction (FDR = 0.05) are as follows: GFP, n = 19, ***p = 1.733 × 10−31; WT UBE3A, n = 19; UBE3A LD, n = 19, ***p = 1.519 × 10−31; R39H, n = 3, p = 0.567; A178T, n = 4, p = 0.828; T485A, n = 5, *p = 0.019; C21Y, n = 3, ***p = 8.725 × 10−6; C117R, n = 3, ***p = 3.419 × 10−4; N243K, n = 3, **p = 0.0072; E269G, n = 3, **p = 7.787 × 10−4; L273F, n = 3, **p = 0.0052; S349P, n = 3, **p = 0.0016; L502P, n = 3, **p = 0.0033; R506C, n = 3, **p = 0.0033; T106K, n = 6, **p = 0.0.0078; T106P, n = 3, **p = 0.0013; I130T, n = 6, *p = 0.016; R477P, n = 3, ***p = 4.24 × 10−4; M478I, n = 3, ***p = 3.39 × 10−4; R482P, n = 3, **p = 5.82 × 10−4; I329T, n = 5, ***p = 1.22 × 10−5; E550L, n = 3, *p = 0.0013. Data reprinted with permission from Weston et al.18. Please click here to view a larger version of this figure.
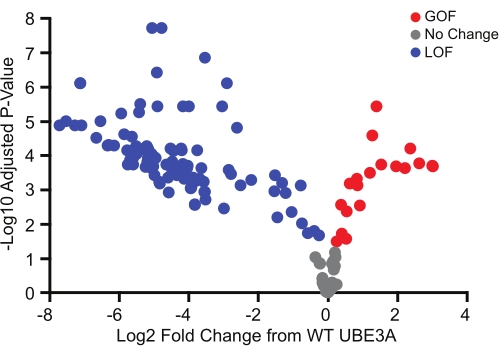
Figure 3: UBE3A variants encompassing a wide range of functional effects. BAR assay screen of 152 Ube3a variants showing benign (gray), loss-of-function (blue), and gain-of-function (red) mutations relative to WT UBE3A. Significance was determined using a one-sample t-test (two-tailed) with Benjamini-Hochberg multiple comparisons correction (FDR = 0.05). Red, gain-of-function; Gray, no change from WT UBE3A; Blue, loss-of-function. Data reprinted with permission from Weston et al.18. Please click here to view a larger version of this figure.
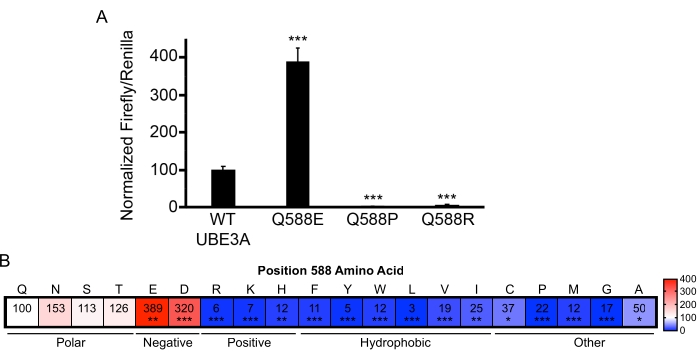
Figure 4: UBE3A variants encompassing a wide range of functional effects. (A) BAR assay results of variants at position 588 of UBE3A showing both loss- and gain-of-function substitutions. Q588E, n = 6; Q588P, n = 3; Q588R, n = 4; ***p < 0.0005, One-sample t-test (two-tailed) with Benjamini-Hochberg multiple comparison correction (FDR = 0.05). (B) Heat plot showing normalized BAR results for comprehensive mutational analysis at position 588 in UBE3A. The white shading represents WT UBE3A activity levels, the blue shading indicates loss-of-function, and the red shading indicates gain-of-function. The scale bar shows the percent change relative to WT UBE3A. N = 3 independent experiments for Q588S, Q588T, Q588N, Q588K, Q588P, Q588M, Q588G, Q588A, Q588F, Q588Y, Q588W, Q588L, Q588V, Q588I, Q588C; n = 8 for Q588E, n = 6 for Q588D, n = 5 for Q588R, n = 4 for Q588H, *p < 0.05, **p < 0.005, ***p < 0.0005. One-sample t-test (two-tailed) with Benjamini-Hochberg multiple comparisons correction (FDR = 0.05). Data reprinted with permission from Weston et al.18. Please click here to view a larger version of this figure.
| Name | Sequence | Used for | |
| Primer 1 | GATTATATTATGACAATAGAA TTCGCATGTACAGTGAAC |
Ube3a EcoRI sense | |
| Mut. Primer | CCAGACCCAGTACTATGCCAAT CAGAGTAAACTCACCCTCAGTT TCAAAAGAAGATGGATTAAACC |
UBE3A Q588E mutagenesis antisense | |
| Primer 2 | ATTATATTCCCGGGTTACAGCAT GCCAAATCCTTTGG |
Ube3a XmaI antisense | |
Table 1: Primers used for UBE3A Q588E mutagenesis.
| 1st Round PCR | |
| Reagent | Reaction Quantity |
| 5x high-fidelity buffer | 10 µL |
| dNTP (10 mM stock) | 1 µL |
| Primer 1 (10 mM stock) | 1 µL |
| Primer 2 (10 mM stock) | 1 µL |
| WT-Ube3a DNA (150 ng/µL stock) | 1 µL |
| H2O | 35 µL |
| High-fidelity DNA polymerase | 1 µL |
| Total Volume | 50 µL |
| 2nd Round PCR | |
| Reagent | Reaction Quantity |
| 5x high-fidelity buffer | 10 µL |
| dNTP (10 mM stock) | 1 µL |
| Primer 3 (10 mM stock) | 1 µL |
| Purified PCR product (Megaprimer) | 30 µL |
| WT-Ube3a DNA (150 ng/µL stock) | 1 µL |
| H2O | 6 µL |
| High-fidelity DNA polymerase | 1 µL |
| Total Volume | 50 µL |
Table 2: PCR parameters for mutagenesis.
| 1st Round PCR | ||
| Step | Temperature | Time |
| Initial denaturation | 95 °C | 2 min |
| (1 cycle) | ||
| Denaturation | 95 °C | 30 s |
| Annealing | 50 °C | 20 s |
| Extension | 72 °C | 15 s |
| (30 cycles) | ||
| Final extension | 72 °C | 1 min |
| Hold | 4 °C | Infinite |
| 2nd Round PCR | ||
| Step | Temperature | Time |
| Initial denaturation | 95 °C | 2 min |
| (1 cycle) | ||
| Denaturation | 95 °C | 30 s |
| Annealing | 50 °C | 20 s |
| Extension | 72 °C | 35 s |
| (30 cycles) | ||
| Final extension | 72 °C | 1 min |
| Hold | 4 °C | Infinite |
Table 3: PCR Program Parameters for Mutagenesis.
| Reagent | Working concentration | 1x transfection | 3.5x master mix |
| pGL3 BAR plasmid | 100 ng/µL | 0.5 µL | 1.75 µL |
| pTK Renilla plasmid | 100 ng/µL | 0.05 µL | 0.175 µL |
| Ube3a plasmid | 100 ng/µL | 0.4 µL | 1.4 µL |
| DMEM supplemented with glutamine | 8.65 µL | 30.275 µL | |
| Transfection reagent | 0.4 µL | 1.4 µL |
Table 4: Transfection mixtures.
