To confirm that 13C6-Glucose supply in rhizomes was successful, we further analyzed the 13C/12C isotope ratios in rhizomes. The 13C /12C isotope ratios of Treatments 3 and 4 were much higher than those of Treatment 2 (Figure 1A). The results indicated that 13C6-Glucose from Treatment 3 and 4 entered the rhizomes through ingestion.
The ratios of 13C isotope peaks, such as (M+1) −/M−, (M+2) −/M−, (M+3) −/M−, and (M+4) −/M−, typically remain constant. Compounds labeled with exogenously added 13C have the same molecular structure, similar molecular weights, physicochemical properties, and retention times as their unlabeled counterparts. However, when these labeled compounds were analyzed using mass spectrometry, the ratios of their isotope ion peak changed. In our study, we used LC-MS to detect PS VII and PS II in four different organs of PPY in various treatments (Figure 3). Compared to Treatment 1, the results indicated a significant increase in the ratio of PS VII (M+2) −/M− in the leaves of the PPY treated with exogenous 13C (Figure 3B). Particularly in Treatment 2, where PPY leaves were sprayed with 13C6-Glucose, PS VII marked with two 13C atoms were synthesized. However, in Treatment 4, the ratio of PS VII molecules (M+2) −/M− in leaves had no significant difference between the rhizome and non-feeding treatment, and the ratios of PS VII molecules (M+1) −/M−, (M+3) −/M−, (M+4) −/M− in stem, rhizome, and root across all four treatments showed no significant differences (Figure 3A,C,D). Additionally, the ratio of PS II molecules (M+2) −/ M− increased slowly in Treatment 2 (Figure 3F). There were no notable differences in the ratios of PS II molecules (M+1) −/M− and (M+4) −/M− in leaves between Treatments 1 and 2 (Figure 3E, H), while the ratio of PS II molecules (M+3) −/M− showed a noticeable decrease in leaves in Treatment 2 compared to Treatment 1 (Figure 3G). These findings suggest that the synthesis of PS VII indeed occurs in the leaves treated with 13C6-Glucose, aligning with the known biosynthetic pathway of PSs.
Our approach suggests the leaf as the primary organ for PS VII synthesis in PPY, contrasting with the synthesis of PS II. Notably, 13C -labeled PS VII was absent in the rhizome following leaf treatment with 13C6-Glucose, implying slow transport of PS VII from leaves to rhizomes. In contrast, 13C -labeled PS II was detected in the rhizome after stem-vascular-bundle and rhizome labeling treatments, indicating possible rhizome synthesis at this stage and necessitating further exploration of PS II synthesis locations. The exclusive detection of the (M+2) −/M− ratio for PS VII molecules in leaves aligns with the metabolic fate of 13C6-Glucose; after absorption, it is metabolized via glycolysis to produce acetyl-CoA with two 13C isotopes. Considering PS as an isoprenoid compound, its biosynthesis from acetyl-CoA proceeds via either the mevalonic acid (MVA) or the 2-C-methyl-D-erythritol-4-phosphate (MEP) pathway (Figure 4), leading to (M+2) −/M− PS VII formation. Thus, our study delineates the leaf as the definitive organ for PS VII synthesis in PPY.
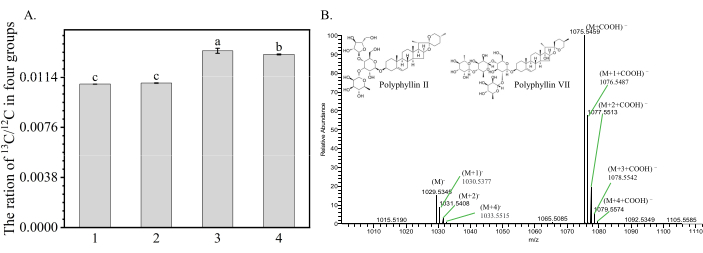
Figure 1: The ratio of 13C /12C isotope in rhizomes of four treatments. (A) The ratios of 13C /12C isotopes in rhizomes of Treatments 4 and 3 were much higher than that of Treatments 1 and 2, indicating that 13C6-Glucose had significantly entered the rhizomes through root feeding. Each column represents the mean (± SE) of three replicates. The letters on the columns mean significantly different in the ratio of 13C/12C isotope in four treatments (p < 0.05). Abbreviation: PS = Paris saponins. (B) The high-resolution mass spectrometry of PS VII (C51H82O21, 1030.5349) detected its negative ion with a retention time of 16.83 minutes. Due to 13C's natural abundance (with an exact molecular weight of 13.003, compared to 12C at 12.000, a range of isotope peaks including (M−), mainly (M+1)−, (M+2)−, (M+3)− and (M+4)−, appear at the same retention time as the PS VII molecular ion peak M−. These peaks were mainly due to COOH− ionization in formic acid water. An enlarged bar diagram highlights these isotope peaks. The ion current ratios of (M+1)−/M−, (M+2)−/M−, (M+3)−/M−, and (M+4)−/M− were calculated. By analyzing changes in these ratios, we could track and infer the biosynthesis of saponins. Here, M− represented the combined peak areas of ion current for (M+COOH)– and M−. Figure 1B was modified from Wen et al.24. Please click here to view a larger version of this figure.
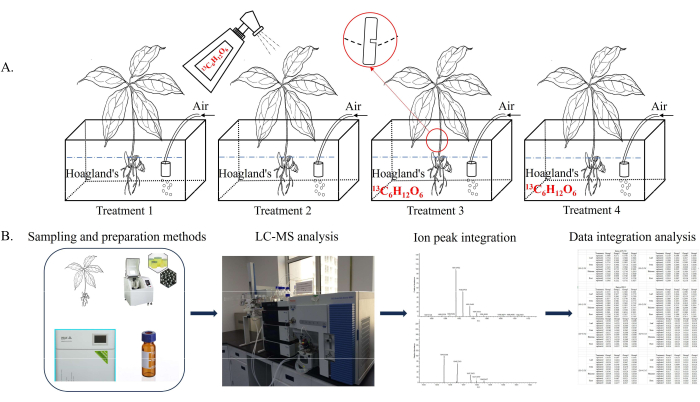
Figure 2: 13C6-Glucose labeling and data Integration workflow. Seedlings were labeled by spraying and feeding 0.2% 13C6-glucose for 3 days; then, extracts from each organ were isotopically analyzed by LC/MS and GC/IRMS and combined with peak integration and data for organ-specific analysis of PS synthesis. (A) A schematic diagram illustrating the feeding of glucose in different organs. Treatments 1-4 were non-feeding, fed in leaf, stem-vascular-bundle, and rhizome, respectively. (B) The steps for LC-MS analysis of the extracted liquid made from the dried PPY tissue through a grinding and nitrogen-blowing instrument. Abbreviations: LC/MS = liquid chromatography-mass spectrometry; PPY = Parispolyphylla var. yunnanensis (Franch.) Hand. -Mzt.; PS = Paris saponin. Please click here to view a larger version of this figure.
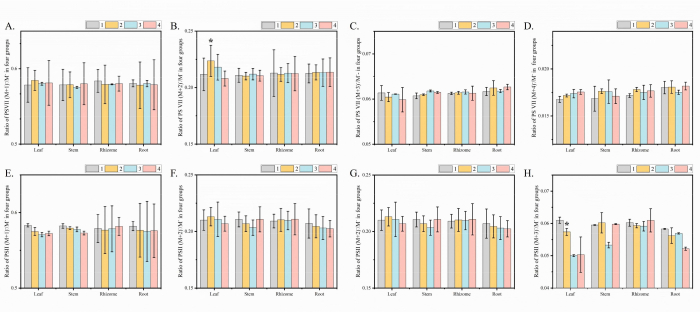
Figure 3: Tracking of Paris saponin VII and Paris saponin II biosynthesis via 13C6-Glucose feeding of plant organs. (A-D) The ratio of PS VII molecules (M+1) −/M−, (M+2) −/M−, (M+3) −/M−, (M+4) −/M− in each organ of the four treatments. The ratio of PS VII molecules (M+2) −/M− increased significantly in the leaves of Treatment 2 and Treatment 3. In the stem, root, and rhizome, the ratio of PS VII molecules (M+2) −/M− showed no obvious differences between the four treatments. (E-H) The ratio of PS VII molecules (M+1) −/M−, (M+2) −/M−, (M+3) −/M−, (M+4) −/M− in each organ of the four treatments. Compared to Treatment 1, the ratios of PS II molecules (M+2) −/ M− in leaves had no difference under Treatment 2. In Treatment 2, the ratio of PS II molecules (M+1) −/M− and (M+4) −/M−have no obvious differences in leaves compared with Treatment 1. The ratios of PS II molecules (M+3) −/M− have decreased in leaves compared with Treatment 1. Paris saponin VII, PS VII. Each column represents the mean (± SE) of three replicates. "*" indicated a significant difference from non-feeding treatment (t-test at p < 0.01). Abbreviation: PS = Paris saponins. Please click here to view a larger version of this figure.
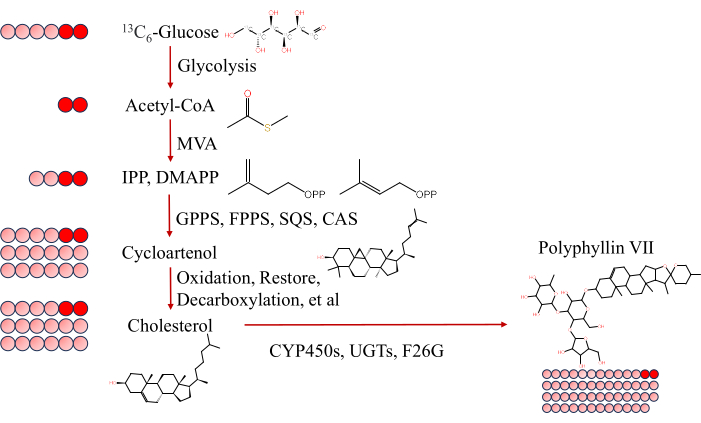
Figure 4: Description of PS VII generated by exogenous 13C6-Glucose entering the MVA pathway. Schematic diagram of the metabolism of 13C6-Glucose-labeled acetyl coenzyme A to generate PS VII molecules (M+2) −/M− via the MVA pathway. Labeled carbon is shown in red; unlabeled carbon is gray. Under the action of glycolysis, 13C6-Glucose is cleaved to two molecules of acetyl coenzyme A(M+2)-, which then generates isopentenyl pyrophosphate and dimethyl propenyl pyrophosphate through the MVA pathway, and then generates cyclic atenol via the catalytic generation of geranylgeranyl pyrophosphate synthase, farnesyl pyrophosphate synthase, squalene synthase, squalene epoxygenase, cycloatenol synthase Cycloartenol, then oxidation, decarboxylation, reduction and other reactions to obtain cholesterol, cholesterol through a series of cytochrome P450 enzymes, glycosyltransferases and furostanoid saponin 26-O-β-glucosidase of the late modification of the formation of PS VII molecules containing two labeled carbon. (M+2) – indicates that two carbon's COOH- is ionized at this point, and the number of labels for two carbon can be derived using (M+2) −/M−. Using standard protocols, the isotopic body is identified individually and the ionic currents were divided to obtain labeling data for the isotopes. The value of M− is stable. Abbreviation: PS = Paris saponins; PS VII = Paris saponin VII; MVA = Mevalonate pathway; GPPS = Geranylgeranyl pyrophosphate synthase; FPPS = Farnesyl pyrophosphate synthase; SQS = Squalene synthase; SQE = Squalene epoxygenase; CYPs = Cytochrome P450 enzymes; UGTs = Glucuronic transferase; F26G = Furostanoid saponin 26-O-β-glucosidase. Please click here to view a larger version of this figure.
