In order to observe the transition from breast non-stem cancer cells to breast CSC-like cells, a first sorting of CD44–/CD24+ breast cancer cells were needed. For the MCF-7 cell line, which has around 0.15% cells with CSC markers in the original population (Figure 1), this step helped exclude the possibility of CSC enrichment during apoptosis reversal. On the contrary, if there were no cells with CSC markers in the original population, such as for T47D cells (Figure 1), this sorting procedure could be omitted. Indeed, gating affected the definition of the positive and negative of each marker, which would eventually influence the percentage of CSC determined. Therefore, appropriate controls including isotype controls for antibodies of interest, single stained controls for each marker should be carefully chosen and prepared for gate adjustment (Figure 1).
With breast non-stem cancer cells, the apoptosis reversal model could thereafter be established. Typical morphological changes could be observed after adding apoptotic inducers and cells should recover from apoptosis with similar morphology after drug withdrawal (Figure 2). Caspase-3/7 recognizes the amino acid sequence DEVD in the Caspase-3/7 Green Detection dye and the active forms of caspase-3/7 are able to cleave this site41. Originally, the fluorescence of the Caspase-3/7 Green Detection dye in cytosol is weak while if the dye is cleaved and translocated to the nucleus, the fluorescence signal would be amplified after its binding to DNA in the nucleus. This obvious difference could be distinguished by flow cytometry. Hence, those caspase-activated cells could be labelled and sorted out based on their higher fluorescence intensity comparing to those without caspase activation (Figures 3A-3D). During the apoptotic induction process, an appropriate solvent control must be included: solvent-treated cells without caspase activation were collected (Figures 3E and 3F) to exclude the possibility that the FACS procedure or the solvent itself was the cause for transition, if any.
In order to show that those FACS-sorted caspase-activated cells were indeed apoptotic, we co-stained these cells with Annexin V and PI. One of the early-stage changes in apoptotic cells is the translocation of phosphatidylserine from the inner side of plasma membrane to the external surface of the cell13,38,39,40; this externalization of phosphatidylserine could be detected by Annexin V. While this change is not unique to apoptosis, PI can be used for distinguishing apoptosis from necrosis based on the integrity of the cell membrane. Thus, cells with Annexin V binding but without PI staining are regarded as apoptotic cells. Caspase-activated cells in the apoptotic inducer treatment groups were found to be Annexin V positive and PI negative, suggesting that they were apoptotic cells (Figure 4).
After the second sorting based on caspase activation in apoptotic cells, these apoptotic cells were collected and subsequently cultured for recovery. The reversed cells that were alive were able to attach the culture container bottom and continue to proliferate. After 7 days, staining of CD44 and CD24 was performed in the reversed cells while controls were prepared at the same time as before. Compared with the solvent-treated groups (Figure 1), flow cytometric analysis showed that there were events (cells) appearing in the CD44+/CD24– quadrant in the reversed breast non-stem cancer cell population (Figure 1). Since we had already excluded cells with CD44+/CD24– in advance of apoptosis induction and only CD44–/CD24+ breast non-stem cancer cells were chosen, these CD44+/CD24– CSC-like cells could only be transited from breast non-stem cancer cells during apoptosis reversal.
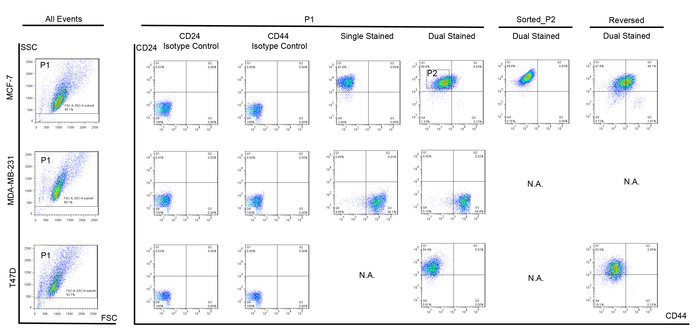
Figure 1: Representative breast CSC marker staining on breast cancer cells in flow cytometry.
MCF-7, MDA-MB-231 and T47D were stained with fluorochrome-conjugated monoclonal antibodies against human CD44 (PerCP-Cy5.5) and CD24 (PE). Cells were first gated by forward scatter (FSC) and side scatter (SSC) (P1) to exclude debris. Isotype controls of CD24 and CD44 were used as negative controls for CD24 and CD44 respectively. MCF-7 cells stained with CD24 was used as positive control for CD24 and MDA-MB-231 cells stained with CD44 was used as positive control for CD44. Non-stem MCF-7 breast cancer cells (P2) were sorted. These sorted cells and T47D cells were subjected to apoptosis reversal procedure. CSC-like (CD44+CD24–) cells appeared after apoptosis reversal. Please click here to view a larger version of this figure.
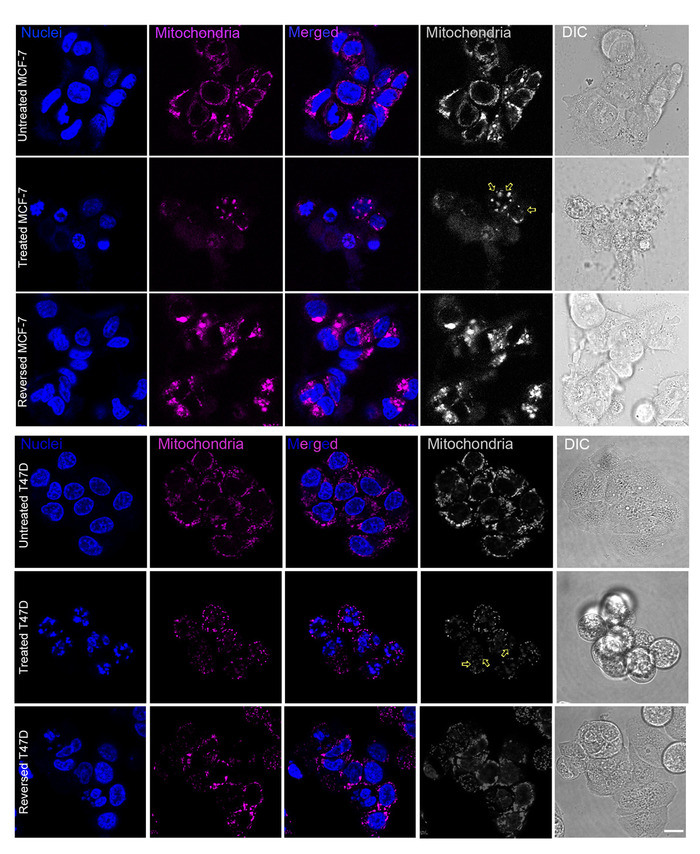
Figure 2: Morphological changes of breast cancer cells under apoptotic stimulus induction.
Breast cancer cells showed typical apoptotic morphological changes including cell shrinkage, membrane blebbing, mitochondria fragmentation (yellow arrows in the monochrome figures) and nuclear condensation. Nuclei (in blue): nuclei of cells stained with Hoechst 33342 were shown in blue color. Mitochondria (in pink): mitochondria of cells stained with Mitotracker Red CMXRos were shown in pink color. Merged: figures merged in dual colors showing both mitochondria and nuclei. Mitochondria (in gray): mitochondria of cells stained with Mitotracker Red CMXRos were shown in monochrome. Differential Interference Contrast (DIC): whole cells. Upper: MCF-7 cells were treated with staurosporine (STS) for 6 h then reversed for 24 h. Lower: T47D cells were treated with paclitaxel for 10 h with reversal for 24 h. Scale bars = 20 μm. Please click here to view a larger version of this figure.
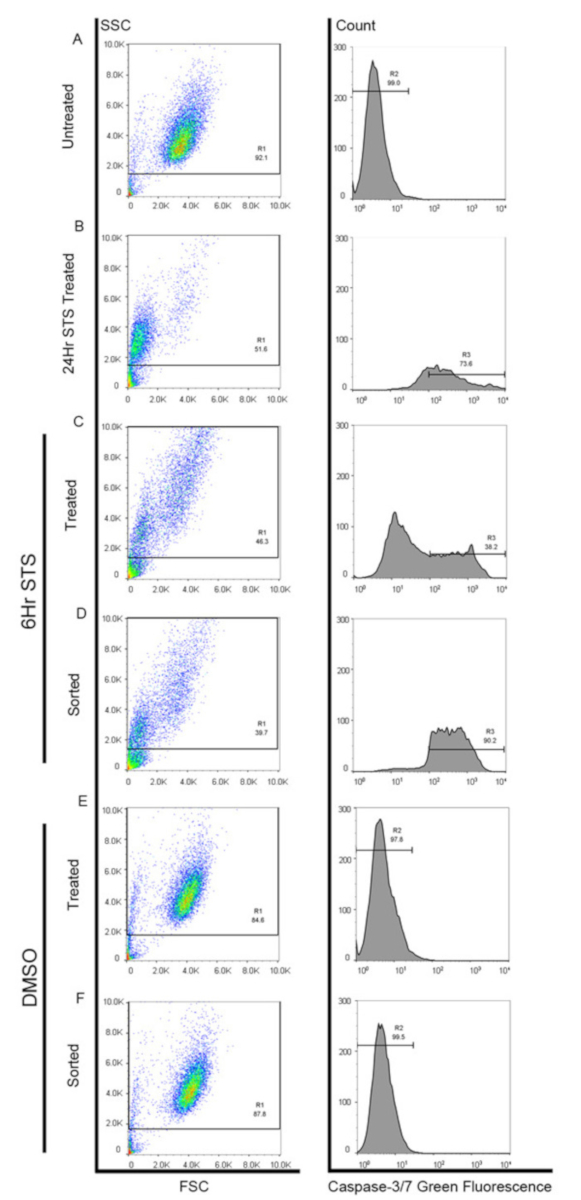
Figure 3: Caspase activation analysis by sorter. (A) Unstained MCF-7 cells without staurosporine (STS) treatment were used as negative control (R2). (B) MCF-7 cells treated with staurosporine (STS) for 24 h. Cells in R3 region were regarded as the caspase-activated cells. (C) MCF-7 cells treated with staurosporine (STS) for 6 h. Cells in R3 region were FACS-sorted. (D) FACS-sorted caspase-activated MCF-7 cells after staurosporine (STS) treatment for 6 h were re-run. (E) DMSO-treated MCF-7 cells. (F) FACS-sorted cells without caspase activation after DMSO treatment for 6 h were re-run. Please click here to view a larger version of this figure.
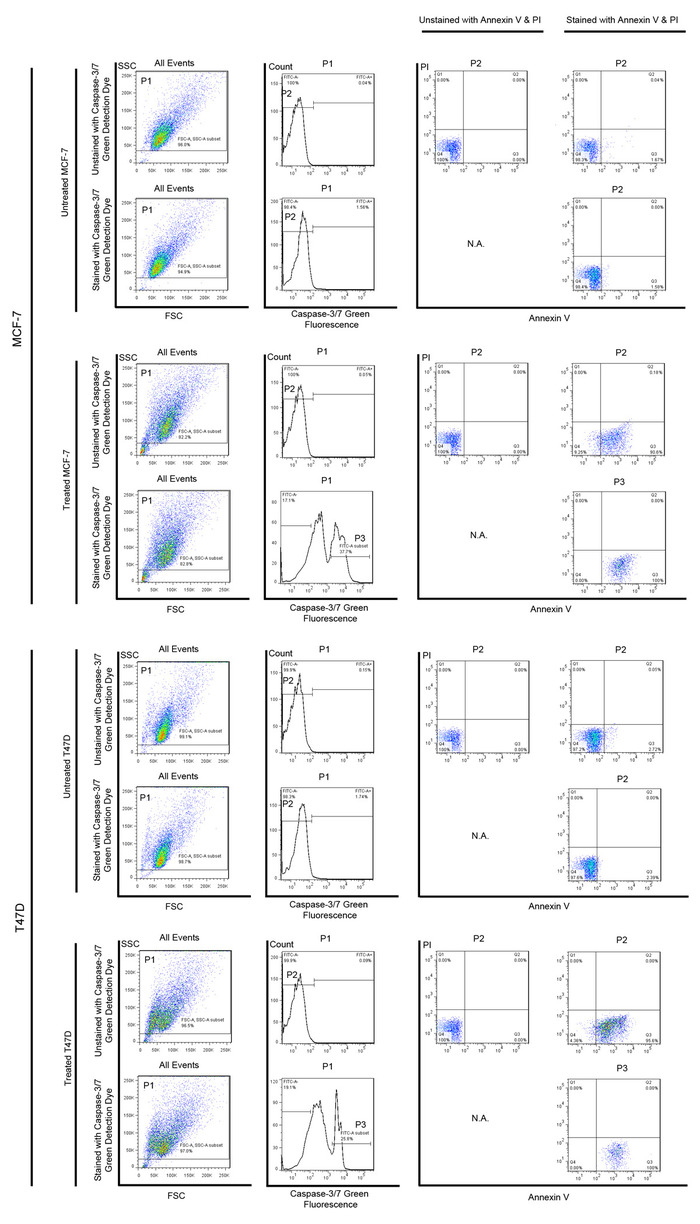
Figure 4: Annexin V and PI staining of caspase-activated cells in flow cytometry. MCF-7 and T47D cells were firstly gated by forward scatter (FSC) and side scatter (SSC) (P1) to exclude debris. Cells that were untreated with apoptotic inducers and without any staining were used as negative controls. For cells treated with apoptotic inducers, caspase-activated cells [i.e., Caspase-3/7 Green positive cells] were selected (P3) to show the fluorescence intensity of Annexin V and PI staining. All Caspase-3/7 Green positive cells were Annexin V positive and PI negative, suggesting that they were apoptotic. Please click here to view a larger version of this figure.
