Клетки человека постоянно подвергаются воздействию различных ДНК-повреждающих веществ различного происхождения. Экзогенные источники в основном состоят из воздействия радиации, химических веществ (включая химиотерапевтические агенты и некоторые антибиотики) и вирусов, в то время как основные эндогенные источники включают ошибки в репликации ДНК и окислительного стресса. Прямые последствия генотоксического воздействия могут варьироваться от модифицированной базы до потенциально смертельного разрыва двойной нити ДНК (DSB), в зависимости от стресса и дозы воздействия. В конечном счете, невосстановимые или неправильно отремонтированные повреждения ДНК могут привести к накоплению мутаций, геномных перестановок, нестабильности генома и в конечном итоге привести кканцерогенеза 1. Клетки млекопитающих развились сложные пути для распознавания конкретных типовповреждения ДНК 2,3 и ремонта их своевременно, синхронизированы с прогрессированием клеточного цикла.
Ионизирующее излучение (ИК) повреждает двойную спираль ДНК и создает двойные разрывы (DSB), одну из самых вредных форм повреждения ДНК. Комплекс MRN (MRE11, RAD50, NBS1) функционирует как датчик конца ДНК и активирует белок киназы атаксия телангиектазия мутировавшая(ATM) 4,5. После первоначальной активации банкомата по ДНК заканчивается, банкомат запускает каскад событий DDR в месте разрыва, инициируя с ключевым событием, фосфорилирование гистона вариант H2AX6. Фосфорилирование H2AX на остатке S139 активирует его в QH2AX, охватывающих регионы до мегабазы вокругпоражения ДНК 6,,7,,8,,9. Это событие повышает доступность ДНК, что приводит к набору и накоплению других белков восстановленияДНК 7. Из-за того, что ЗГ2АКС обильно и специально индуцирован окружающими DSB, его можно легко визуализировать с помощью специфических антител, и он обычно используется в качестве суррогатного маркера для DSBs в области восстановления ДНК. Как только разрыв сигнализируется, клетки активируют пути восстановления ДНК и обработать повреждение ДНК. Белок MDC1 (посредник белка контрольно-пропускного пункта повреждения ДНК 1) напрямую связывает qH2AX10,взаимодействуетс банкоматом 11, а также с NBS112,,13. Это способствует увеличению концентрации комплекса MRN в DSB и инициированию положительного цикла обратной связи банкоматов. После ремонта разрыв быстро удаляется, что позволяет контролировать клиренс DSB. Вслед за микроскопией, снижение йH2AX с течением времени обеспечивает косвенное измерение остаточных разрывов и эффективности восстановления ДНК.
Эукариотические клетки могут ремонтировать DSBs несколькими путями, два основных из которых являются неомологическими конца присоединения (NHEJ) и гомологичной рекомбинации (HR) (Рисунок 1). NHEJ по существу лигаты ДНК двойной пряди заканчивается без использования расширенной гомологии и работает на протяжении всегоклеточного цикла 14,15. HR становится преобладающим во время фаз S и G2, и в противном случае подавляется, так как он требует сестра хроматид в качестве гомологовогошаблона для ремонта 14,16. Выбор пути между NHEJ и HR зависит не только от физической близости сестры хроматид, но и от расширения ДНК конца ресекции17, который подавляет NHEJ.
Гомологиологически-зависимый ремонт DSB инициирует нуклеолитической деградацией 5′ нитей от концов разрыва для генерации 3′ однотяговых ДНК (ssDNA) хвостов, процесс, называемый 5′-3′ resection. Комплекс MRN инициирует ресекцию конца ДНК и дальнейшая ресекция обрабатывается в сочетании с BLM/EXO1 (белок синдрома Блума/экзонуклеаза 1) или BLM/DNA2 (репликация ДНК АТФ-зависимого геликайса/нуклеазы)18,19,20,21,22. Ресекция конца ДНК усиливается CtIP (CtBP-взаимодействующий белок) через его прямое взаимодействие с MRNкомплекс 23 и набор BRCA1 (белок восприимчивости рака молочной железы типа1) 24,25. Репликационный белок A (RPA) быстро связывается с ssDNA подвергается, а затем вытесняется рекомбиназы белка RAD51 сформировать нуклеопротеин нить, которая катализа гомологичный поиск и вторжениепряди 26,27,28.
Инициирование ресекции является важным шагом для выбора пути ремонта. После того, как ресекция началась, концы ДНК становятся бедными субстратами для связывания гетенодимером Ku70/Ku80 (компонент пути NHEJ), а клетки привержены HR17,,29,30. Ku70/Ku80 гетенодимер связывается с концами DSB, вербовка ДНК-PKcs и p53 Связывающий белок 1 (53BP1)29,30. 53BP1 выступает в качестве барьера для resection в G1, тем самым блокируя HR при продвижении NHEJ31,32, но он удаляется в BRCA1-зависимой манере в фазе S, следовательно, позволяя resectionпроизойти 33,34. Таким образом, 53BP1 и BRCA1 играют противоположную роль в ремонте DSB, с 53BP1 является посредником NHEJ в то время как BRCA1 действует позволяет перерывы для ремонта через HR.
В лаборатории образование DSB может быть вызвано ионизирующим излучением (ИК). Хотя этот пример использует высокую дозу 4 Gy, 1 Gy и 2 Gy также создать значительное количество DSBs, подходит для анализа образования очагов обильными белками. Важно отметить, что тип и доза используемого излучения может привести к различным поражениям в ДНК и в клетке: в то время как ИК индуцирует DSBs, он также может вызвать одиночные разрывы нити или изменения базы(см. 35,36 для справки об облучении линейной передачи энергии (LET) и тип повреждения ДНК). Для определения кинетики ионизирующего радиационно-индуцированного образования очагов (IRIF) и их расчистки, которые указывают на ремонт повреждений и разворот активированного DDR8,,9,,37,,38,образование очагов можно контролировать в разные моменты времени после ионизирующего излучения. Сроки активации и очистки всех основных белков поврежденияДНК известно 39, и многие из них расследуются как суррогатные маркеры ключевых событий. Например, pRPA, который обладает высокой сродством к ssDNA используется в качестве суррогата ресекции разрыва, белки MRN (MRE11, RAD50, NBS1) и экзонуклеазы могут быть использованы для оценки эффективности ресекции тоже. В то время как RAD51, BRCA1, BRCA2 (белок восприимчивости к раку молочной железы 2 типа) и PALB2 (партнер и локализатор BRCA2) могут быть проверены для оценки эффективности HR, наличие белков Ku или 53BP1, используются в качестве маркеров NHEJ (Рисунок 1).
По мере того как протеины машин ремонта ДНАА вербуют один другого к пролому и собирают в супер-комплексах, взаимодействия дна-белка и протеина-протеина можно сделать вывод путем следовать за их индивидуальной локализацией над временем и анализировать co-локализацию протеинов, как визуализировано перекрывая сигналамив клетке 40,41,42. В клеточных линиях введение точечных мутаций или удаление в конкретных генах восстановления ДНК либо путем редактирования генома, либо путем переэкспрессии плазмидных мутантов, позволяет изутовить специфические остатки и их возможную роль в признании повреждения ДНК (например, совместной локализации с помощью QH2AX) или сложной сборки (совместной локализации с другим, или несколькими белками), а также их влияния на ДНК. Здесь мы используем косвенное иммунофторесценцию как среднее для исследования образования и разрешения DSBs, следуя фоклюсу NO2AX с течением времени. Мы также представляем один пример анализа формирования очагов и совместной локализации крупным игроком в ремонте DSB: p53 Binding Protein 1 (53BP1)32. Как упоминалось ранее, 53BP1 считается центральным для выбора пути восстановления ДНК. После накопления 53BP1 и его совместной локализации с хХ2АКС предоставляет ценную информацию о фазе клеточного цикла, накоплении повреждений ДНК и пути, используемом для ремонта DSB. Целью косвенной иммунолокализации является оценка эффективности восстановления повреждений ДНК в клеточных линиях, после ИК, как в данном исследовании, или после воздействия различных напряжений в клетке, от перекрестного пересечения ДНК до блокировки вилки репликации (список повреждающих ДНК агентов приведен в таблице 1).
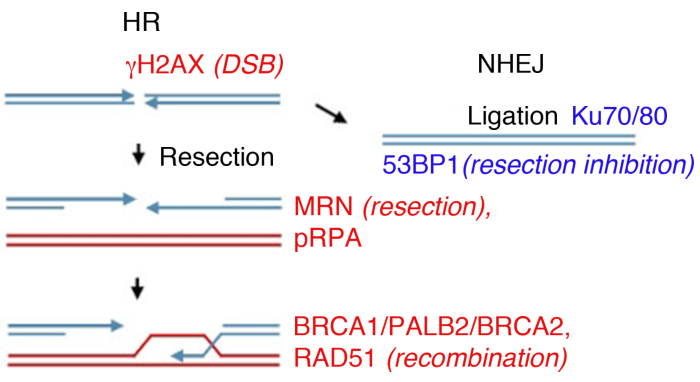
Рисунок 1: ДНК двойные нити перерывов (DSB) ремонт путей.
Ремонт DSB включает в себя два основных пути: гомологичная рекомбинация (HR, слева) и не гомологичный end-Joining (NHEJ, справа). После перерыва, белки активируются, чтобы отметить разрыв (NOH2AX), участвовать в конце ресекции (MRN), пальто resected ssDNA (pRPA), способствовать рекомбинации (BRCA1, PALB2, BRCA2, RAD51) или ограничить ресекцию и содействовать NHEJ (53BP1). Другие белки участвуют в ремонте повреждений, но перечисленные белки обычно следуют косвенные иммунофлуоресценции. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
| Повреждающее ДНК агент | Механизм действия | Рекомендуемая доза |
| К-лучи/рентгеновские лучи | Излучения Формирование двухместных разрывов с некоторыми неконтролируемыми клеточными эффектами |
1-4 Gy |
| 36 Лет Ар ионы | Излучения Формирование двухместных перерывов |
270 кВ/мкм |
| К-частицы | Излучения Формирование двухместных перерывов |
116 кВ/мкм |
| Блеомицин | Ингибитор синтеза ДНК | 0,4-2 мкг/мл |
| Камптотецин | Ингибитор топоизомеразы I | 10-200 нм |
| Цисплатин | Алкилайт агент (индуцирующие внутристрандовые перекрестные ссылки) |
0,25-2 МКМ |
| Доксорубицин | Интеркалирующий агент Ингибитор топоизомеразы II |
10-200 нм |
| Этопозид | Ингибитор топоизомеразы II | 10 МКМ |
| Гидроксиуреа | Ингибитор синтеза ДНК (по рибонуклеотидной редуказе) |
10-200 МКМ |
| Метил метансульфонат | Алкилайт агент | 0,25-2 мММ |
| Митомицин C | Алкилайт агент | 0,25-2 МКМ |
| Ультрафиолетовый (УФ) свет | Формирование димеров тимидин (генерация искажения цепочки ДНК) |
50-100 мДж/см2 |
Таблица 1: Генотоксические агенты. Примеры повреждающих ДНК агентов, их механизм действия и ущерб, индуцированный на основе предложенной рабочей концентрации.
