As células imunes defendem-se contra patógenos e células cancerígenas rastejando e varrendo continuamente as superfícies das células-alvo em busca de antígenos, fixando sua superfície 1,2. O reconhecimento do antígeno é iniciado após a ligação entre o receptor de células T (TCR) e o complexo MHC (pMHC) do complexo peptídeo-principal de histocompatibilidade expresso na superfície das células-alvo. Como o reconhecimento do TCR-pMHC ocorre na junção entre duas células móveis, há muito se suspeita que ele experimente forças mecânicas. Além disso, isso levou ao modelo mecanosensor de ativação do TCR, que sugere que as forças do TCR contribuem para sua função 3,4. Para entender quando, onde e como as forças mecânicas contribuem para o funcionamento das células T, é imperativo desenvolver ferramentas para visualizar as forças moleculares transmitidas pelas células T. Tradicionalmente, métodos como a microscopia de força de tração (MFT) e os arranjos de micropilares são utilizados para investigar as forças celulares 5,6. No entanto, a sensibilidade à força dos arranjos de MTF e micropilares está na escala de nanonewton (nN) e, portanto, muitas vezes é insuficiente para estudar as forças moleculares de piconewton (pN) transmitidas por receptores celulares7. Para melhorar a força e a resolução espacial para detecção, nosso laboratório foi pioneiro no desenvolvimento de sondas de tensão molecular, que foram inicialmente sintetizadas usando polímeros de polietilenoglicol (PEG)7. As sondas de tensão molecular são compostas por uma “mola” molecular extensível (PEG, proteína, DNA) flanqueada por um fluoróforo e quencher e estão ancoradas em uma superfície. Forças aplicadas ao término da sonda levam à sua extensão, separando o fluoróforo e o quencher, gerando um forte sinal de fluorescência (Figura 1A)8,9,10.
Na última década, desenvolvemos uma biblioteca de diferentes classes de sondas de tensão molecular com elementos de mola feitos de ácidos nucléicos11, proteínas10 e polímeros8. Dentre estas, as sondas de tensão baseadas em DNA fornecem a maior relação sinal/ruído e a maior sensibilidade à força, que é facilmente sintonizada de alguns pN até ~20 pN11. Usamos essas sondas de tensão de DNA em tempo real para estudar as forças moleculares geradas por diversos tipos celulares, incluindo fibroblastos, células cancerosas, plaquetas e células imunes11,12,13. Este manuscrito descreverá protocolos para sintetizar e montar sondas de tensão de DNA em uma superfície para mapear forças de receptores moleculares com resolução de força de pN usando um microscópio de fluorescência convencional. Embora o procedimento atual inclua modificações químicas no ácido nucleico para introduzir o repórter fluorescente (Figura 1B), é importante notar que muitas das etapas de modificação e purificação podem ser terceirizadas para empresas de síntese de DNA personalizadas. Portanto, a tecnologia de sondas de tensão de DNA é fácil e acessível para as comunidades mais amplas de biologia celular e mecanobiologia.
Resumidamente, para montar sensores de tensão de DNA, um hairpin de DNA é hibridizado a uma fita fluorescente ligante em um braço e uma fita de âncora quencher no outro braço e, em seguida, imobilizado em um substrato de vidro (Figura 1C, tensão em tempo real). Na ausência de força mecânica, o hairpin é fechado e, assim, a fluorescência é apagada. No entanto, quando a força mecânica aplicada é maior que o F1/2 (a força em equilíbrio que leva a uma probabilidade de 50% de desdobramento), o hairpin derrete mecanicamente e um sinal fluorescente é gerado.
Com base no sensor de tensão do DNA em tempo real, também descrevemos protocolos para mapear forças acumuladas, o que é particularmente útil para estudar interações entre receptores em células imunes e seu ligante natural. Isso ocorre porque os receptores imunes frequentemente apresentam ligações de curta duração 3,14. As forças acumuladas são imageadas usando uma fita de “travamento” que se liga preferencialmente aos pinos de cabelo de DNA abertos e permite o armazenamento de sinais de fluorescência associados a eventos de tração mecânica (Figura 1C, tensão bloqueada). O fio de bloqueio é projetado para ligar um local de ligação críptico que é exposto após o derretimento mecanicamente induzido do hairpin e travar o hairpin no estado aberto, bloqueando a redobração do hairpin, armazenando assim o sinal de tensão e gerando um mapa de tensão acumulado. Além disso, a fita de travamento é projetada com um suporte de oito nucleotídeos, que permite uma reação de deslocamento de fita mediada por dedos dos pés com seu complemento completo, a fita de “destravamento”. Com a adição do fio de desbloqueio, o fio de travamento preso é retirado da construção do hairpin, apagando o sinal de tensão armazenado e redefinindo o hairpin de volta ao estado em tempo real.
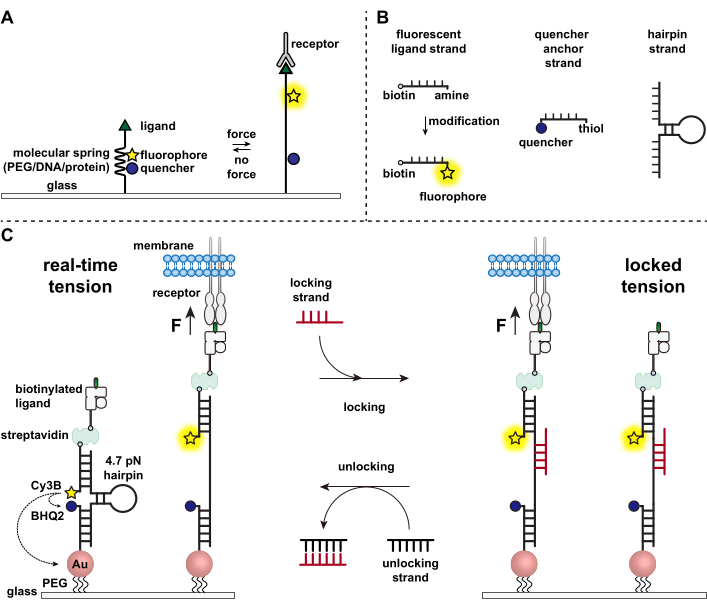
Figura 1: Esquema das sondas de tensão molecular de última geração. (A) Projeto geral de sonda de tensão molecular em tempo real, (B) Fitas para a construção de sonda de tensão baseada em DNA e (C) Sondas de tensão baseadas em DNA projetadas e sua alternância entre o estado em tempo real e o estado bloqueado. Clique aqui para ver uma versão maior desta figura.
O protocolo principal consiste em quatro seções principais – preparação de oligonucleotídeos, preparação de superfície, imagem e análise de dados. Este protocolo foi demonstrado com sucesso por nosso laboratório e outros em células T CD8+ OT-1 virgens e ativadas, células CD4+ OT-II, bem como hibridomas, e pode ser aplicado para interrogar diferentes receptores de células imunes, incluindo receptor de células T, receptor de morte celular programada (PD1) e forças do antígeno 1 associado à função linfocitária (LFA-1). Células T virgens de OT-1 CD8+ são usadas como um exemplo de linhagem celular neste artigo.
