카스파제는 개시제 카스파제 및 집행자 카스파제로 그룹화될 수 있는 시스테인 아스파르테이트 프로테아제의 패밀리이다. 사형 집행자 카스파제는 카스파제-3, -6 및 -7을 포함한다. 그들은 이량체로서 세포에서 자연적으로 발견되며 아폽토시스1을 실행하기 위해 개시제 카스파제에 의해 절단됩니다. 개시제 카스파제는 인간 카스파제-1, -2, -4, -5, -8, -9, -10 및 -12를 포함한다. 이들은 근접-유도된 이량체화에 의해 활성화되고 자가-단백질 분해 절단 2,3에 의해 안정화되는 비활성 효소원(pro-caspases)으로서 발견된다. 염증성 카스파제는 개시제 카스파제2의 서브세트이고, 인간에서 카스파제-1, -4, -5, 및 -12를 포괄하고, 마우스4,5에서 카스파제-1, -11, 및 -12를 포괄한다. 아폽토시스 역할보다는 염증에서 중심적인 역할을합니다. 이들은 프로인터루킨(IL)-1β 및 프로-IL-18 6,7의 단백질 분해 처리 및 분비를 매개하며, 이는 병원성 침입자 8,9에 반응하여 방출되는 최초의 사이토카인이다. Caspase-1은 활성화 플랫폼으로의 모집시 활성화됩니다. 큰 분자량의 단백질 복합체를 인플람마솜(도 1A)10이라고 불렀다. 카스파제-4, -5 및 -11의 이량체화는 비정준 인플라마솜 경로(11,12)를 통해 이들 플랫폼과 독립적으로 발생한다.
정준 인플라마좀은 인플람마솜 센서 단백질, 어댑터 단백질 ASC(CARD를 함유하는 아폽토시스-관련 얼룩-유사 단백질), 및 이펙터 단백질 카스파제-110으로 구성된 시토졸 다량체 단백질 복합체이다. 가장 잘 연구된 정준 인플라마좀은 피린 도메인 (NLRP), NLRP1 및 NLRP3을 함유하는 NOD 유사 수용체 패밀리, CARD (NLRC), NLRC4, 및 흑색종 2 (AIM2)에 결석한 NLR 패밀리를 함유한다. 그들은 각각 피린 도메인, 카드 또는 두 도메인을 모두 포함합니다. CARD 도메인은 CARD 함유 카스파제와 그들의 업스트림 활성화제 사이의 상호작용을 매개한다. 따라서, N 말단 피린 도메인 (PYD) 및 C 말단 CARD 모티프 13,14로 구성되는 스캐폴드 분자 ASC는 NLRP1 10, NLRP315 및 AIM216 inflammasomes에 대한 카스파제-1의 모집에 필요하다.
각 인플람마솜은 뚜렷한 전염증성 자극을 인식하는 고유한 센서 단백질의 이름을 따서 명명되었습니다(그림 1B). 이 경로의 활성화자는 정식 자극이라고합니다. 인플람마좀은 미생물 성분 및 조직 스트레스에 대한 센서로서 작용하고, 염증 카스파제(17)의 활성화를 통해 강력한 염증 반응을 촉발시키기 위해 조립된다. 인플라마솜 어셈블리는 카스파제-1 활성화를 개시하여 그의 주요 기질인 pro-IL-1β 및 pro-IL-18의 성숙 및 분비를 매개한다. 이 프로세스는 두 단계 메커니즘을 통해 발생합니다. 첫째, 프라이밍 자극은 NF-κB 경로의 활성화를 통해 특정 염증성 단백질 및 pro-IL-1β의 발현을 상향조절한다. 둘째, 세포내(정경적) 자극은 프로파스파제-1 6,7의 인플라마솜 조립 및 모집을 유도한다.
Caspase-4 및 caspase-5는 뮤린 카스파제-11 11의 인간 오르토로그이다. 이들은 그람 음성 박테리아 18,19,20의 외막에서 발견되는 분자인 세포내 리포폴리사카라이드(LPS)와 적혈구 용혈(21)의 산물인 세포외 헴에 의해 인플라마솜-비의존성 방식으로 활성화된다. LPS가 이들 단백질의 CARD 모티프에 직접 결합하고 이들의 올리고머화(20)를 유도한다는 것이 제안되었다. 카스파제-4 또는 카스파제-5의 활성화는 기공 형성 단백질 가스더미민 D(GSDMD)18,19의 절단을 통해 피롭토시스라고 불리는 염증성 형태의 세포 사멸을 유도함으로써 IL-1β 방출을 촉진한다. 또한, 카스파제-4 및 GSDMD 매개 열록토시스 사멸로부터 야기되는 칼륨 이온의 유출은 NLRP3 인플라마좀의 활성화 및 카스파제-122,23의 후속 활성화를 유도한다. 따라서, 카스파제-4, -5, 및 -11은특정 자극(11,24)에 반응하여 피옵토시스 및 카스파제-1 활성화를 유도할 수 있는 LPS용 세포내 센서로 간주된다.
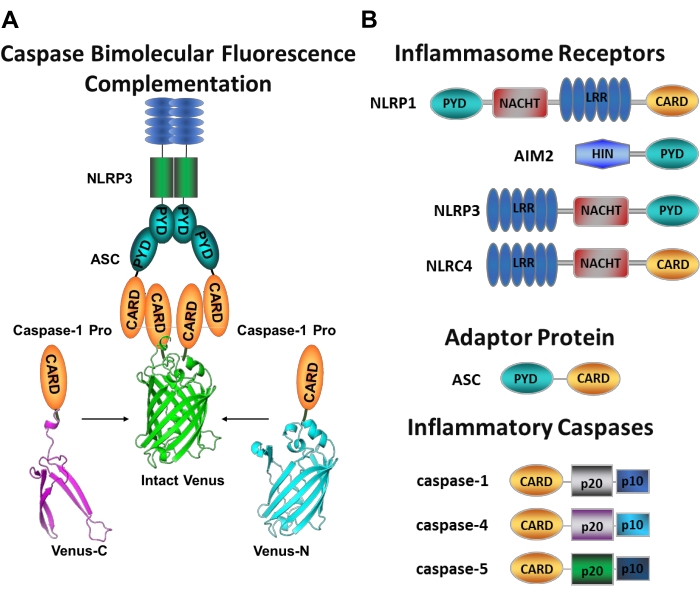
그림 1: 염증성 카스파제 및 카스파제-이분자 형광 보체 (BiFC) 분석. (A) 카스파제-BiFC 시스템을 나타낸 도면으로, 금성의 각 비형광 단편(Venus-C 또는 Venus-N)에 연결된 두 개의 카스파제-1 프로도메인(C1-pro)이 NLRP3 활성화 플랫폼으로 모집되어 금성이 재굴 및 형광을 갖도록 강제한다. 이 복합체는 현미경 하에서 녹색 반점으로 나타나며 개시제 카스파제 활성화의 첫 번째 단계 인 염증성 카스파제 유도 근접성에 대한 판독 역할을합니다. (b) 염증성 카스파아제 및 염증성 카스파제의 도메인 조직을 보여주는 개략도. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
특정 개시제 카스파제 활성화를 측정하는 것은 어렵고, 이미징 접근법에 의해 그렇게 할 수 있는 방법은 많지 않다. Caspase Bimolecular Fluorescence Complementation (BiFC)은 살아있는 세포에서 직접 염증성 카스파제 활성화를 시각화하는 데 사용할 수 있습니다 (그림 1A)25. 이 기술은 최근 인간 단핵구 유래 대식세포 (MDM)21에서 사용하기 위해 적응되었다. 카스파제 BiFC는 염증성 카스파제 활성화의 첫 번째 단계를 측정하고, 이량체화를 촉진하기 위해 근접성을 유도한다. 광화성 황색 형광 단백질 금성 (Venus-C[VC]) 및 Venus-N[VN])의 비형광 단편에 융합된 CARD 함유 카스파제 프로도메인을 코딩하는 플라스미드의 발현이 사용된다. 두 카스파제 프로도메인이 그들의 활성화 플랫폼으로 모집되거나 유도된 근접성을 겪을 때, 금성의 두 반쪽은 근접하게 가져와 굴착 및 형광을 강요받는다(도 1A,B 참조). 이는 특정 염증성 카스파제 활성화의 실시간 판독을 제공한다.
인간 MDM은 위험 신호 및 병원체 생성물을 식별하는 인플라마솜 유전자와 패턴 인식 수용체를 풍부하게 발현한다. 이것은 염증성 카스파제 경로의 심문을 위한 이상적인 세포 유형을 제공한다. 또한, 이들은 말초 혈액 및 심지어 환자 샘플로부터 유래되어 특정 질병 상태에서의 염증성 카스파제 활성화를 평가할 수 있다. 이 프로토콜은 뉴클레오펙션을 사용하여 BiFC 카스파제 리포터를 MDM에 도입하는 방법, 전기천공 기반 트랜스펙션 방법, 염증성 카스파제 활성화를 유도하기 위해 세포를 치료하는 방법, 현미경 접근 방식을 사용하여 활성 카스파제 복합체를 시각화하는 방법을 설명합니다. 추가적으로, 이 방법론은 이들 복합체의 분자 조성, 아세포 국재화, 동역학, 및 이들 고도로 정렬된 구조의 크기(25,26,27)를 결정하도록 적응될 수 있다.
