Der menschliche Körper beherbergt schätzungsweise 10-100 Billionen lebende mikrobielle Zellen (Bakterien, Archaeenpilze), die hauptsächlich in der Darm-, Haut- und Schleimhautumgebung vorkommen1. In einem gesunden Zustand bieten diese ihrem Wirt Vorteile, einschließlich Vitaminproduktion, Reifung des Immunsystems, Stimulation angeborener und adaptiver Immunantworten auf Krankheitserreger, Regulierung des Fettstoffwechsels, Modulation von Stressreaktionen und mehr, mit Auswirkungen auf Wachstum und Entwicklung, Krankheitsbeginn und Alterung 2,3,4,5 . Die Darmmikrobiota entwickelt sich auch im Laufe des Lebens erheblich. Die drastischste Entwicklung findet im Säuglingsalter und in der frühen Kindheit statt6, aber auch mit zunehmendem Alter treten signifikante Veränderungen auf, einschließlich einer Abnahme der Bifidobacterium-Häufigkeit und einer Zunahme von Clostridium, Lactobacillus, Enterobacteriaceae und Enterococcus-Arten 7. Der Lebensstil kann die mikrobielle Zusammensetzung des Darms weiter verändern, was zu Dysbiose führt (Verlust nützlicher Bakterien, übermäßiges Wachstum opportunistischer Bakterien), was zu verschiedenen Pathologien wie entzündlichen Darmerkrankungen, Diabetes und Fettleibigkeit führt5, aber auch zu Alzheimer und Parkinson beiträgt 8,9,10,11.
Diese Erkenntnis hat entscheidend dazu beigetragen, das Konzept der Darm-Hirn-Achse (GBA) zu verfeinern, bei der Wechselwirkungen zwischen der Darmphysiologie (die jetzt die darin enthaltenen Mikroben einschließt) und dem Nervensystem als Hauptregulator des tierischen Stoffwechsels und der physiologischen Funktionen angesehen werden12. Die genaue Rolle der Mikrobiota bei der Darm-Hirn-Signalübertragung und die damit verbundenen Wirkmechanismen sind jedoch noch lange nicht vollständig verstanden13. Da die Darmmikrobiota eine Schlüsseldeterminante für gesundes Altern ist, ist die Art und Weise, wie Bakterien den Alterungsprozess modulieren, Gegenstand intensiver Forschung und Kontroversen geworden 6,14,15.
Mit dem Nachweis, dass der Spulwurm Caenorhabditis elegans eine bonafide Darmmikrobiota beherbergt, die – wie bei anderen Arten – von Bacteroidetes, Firmicutes und Actinobacteria dominiert wird 16,17,18,19,20, sein rasanter Aufstieg als experimentelle Plattform zur Untersuchung kommensaler Interaktionen zwischen Wirt und Darm21,22,23,24 ,25,26 hat unser Ermittlungsarsenal 26,27,28,29 erheblich erweitert. Insbesondere Hochdurchsatz-experimentelle Ansätze, die für C. elegans zur Verfügung stehen, um Gen-Diät-, Gen-Medikament-, Gen-Pathogen-Interaktionen usw. zu untersuchen, können angepasst werden, um schnell zu untersuchen, wie bakterielle Isolate und Cocktails die Gesundheit und das Altern von C. elegans beeinflussen.
Das vorliegende Protokoll beschreibt eine experimentelle Pipeline, um sofort Arrays von Bakterienisolaten oder Mischungen in Multiwell-Platten auf Auswirkungen auf die Stressresistenz von C. elegans als Proxy für die Gesundheit zu untersuchen, die zur Identifizierung von Probiotika verwendet werden können. Es wird detailliert beschrieben, wie große Wurmpopulationen gezüchtet und bakterielle Arrays in 96- und 384-Well-Plattenformaten gehandhabt werden, bevor Würmer für die automatisierte Stressresistenzanalyse mit einem Fluoreszenzplattenleser verarbeitet werden (Abbildung 1). Der Ansatz basiert auf markierungsfreien automatisierten Überlebensassays (LFASS)30, die das Phänomen der Todesfluoreszenz31 ausnutzen, wobei sterbende Würmer einen Ausbruch blauer Fluoreszenz erzeugen, mit dem der Zeitpunkt des Todes bestimmt werden kann. Blaue Fluoreszenz wird von Glucosylestern der Anthranilsäure emittiert, die in C. elegans-Darmgranulaten (einer Art Lysosomen-verwandter Organelle) gespeichert sind, die platzen, wenn eine nekrotische Kaskade im Wurmdarm nach dem Tod ausgelöstwird 31.
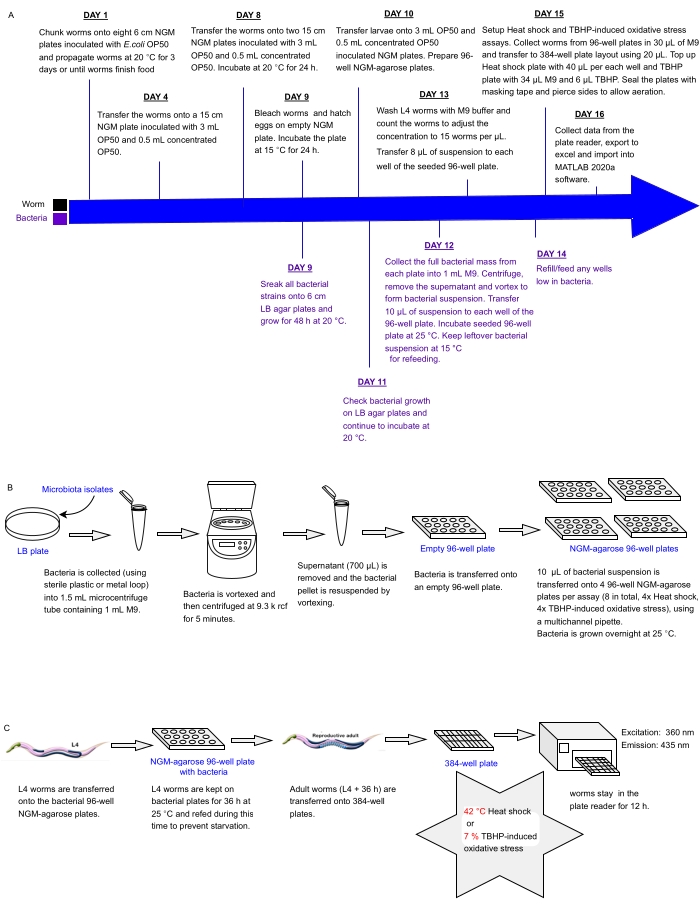
Abbildung 1: Experimenteller Workflow für das Hochdurchsatz-Screening von Bakterienisolaten mit Auswirkungen auf die Stressresistenz von C. elegans . (A) Zeitplan für die Erhaltung von Würmern und Bakterien und den Aufbau des Assays. (B) 96-Well-Aufbau und Handhabung bakterieller Plattenarrays. (C) 384-Well-Schneckenplatten-Setup. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
