El campo de la biología sintética ascendente se centra en la construcción de células (mínimas) 1,2 y biorreactores metabólicos con fines biotecnológicos 3,4 o biomédicos 5,6,7,8. La construcción de células sintéticas proporciona una plataforma única que permite a los investigadores estudiar proteínas (de membrana) en condiciones bien definidas que imitan las de los entornos nativos, lo que permite el descubrimiento de propiedades emergentes y funciones bioquímicas ocultas de las proteínas y las redes de reacción9. Como paso intermedio hacia una célula sintética que funcione de forma autónoma, se desarrollan módulos que capturan características esenciales de las células vivas, como la conservación de la energía metabólica, la síntesis de proteínas y lípidos y la homeostasis. Estos módulos no solo mejoran nuestra comprensión de la vida, sino que también tienen aplicaciones potenciales en los campos de la medicina8 y la biotecnología10.
Las proteínas transmembrana están en el corazón de prácticamente cualquier red metabólica, ya que transportan moléculas dentro o fuera de la célula, señalizan y responden a la calidad del medio ambiente y desempeñan numerosas funciones biosintéticas. Así, la ingeniería de módulos metabólicos en células sintéticas requiere en la mayoría de los casos la reconstitución de proteínas de membrana integrales y/o periféricas en una bicapa de membrana compuesta por lípidos específicos y de alta integridad (baja permeabilidad). El manejo de estas proteínas de membrana es un desafío y requiere conocimientos específicos y habilidades experimentales.
Se han desarrollado varios métodos para reconstituir proteínas de membrana dentro de vesículas de fosfolípidos, la mayoría de las veces con el propósito de estudiar la función11,12, la regulación13, las propiedades cinéticas14,15, la dependencia lipídica15,16 y/o la estabilidad17 de una proteína específica. Estos métodos implican la dilución rápida de proteínas solubilizadas con detergente en medios acuosos en presencia de lípidos18, la eliminación de detergentes mediante la incubación de proteínas solubilizadas con detergente con vesículas lipídicas desestabilizadas con detergente y la absorción de los detergentes en perlas de poliestireno19, o la eliminación de detergentes mediante diálisis o cromatografía de exclusión por tamaño20. Los disolventes orgánicos se han utilizado para formar vesículas lipídicas, por ejemplo, a través de la formación de interfases aceite-agua21, pero la mayoría de las proteínas integrales de la membrana se inactivan cuando se exponen a dichos disolventes.
En nuestro laboratorio, reconstituimos principalmente proteínas de membrana por el método de absorción de detergente para formar vesículas unilaminares grandes (LUVs)19. Este método permite la co-reconstitución de múltiples proteínas de membrana y la encapsulación en la luz de la vesícula de enzimas, metabolitos y sondas22,23. Las LUV que contienen proteínas de membrana pueden convertirse en vesículas unilamelares gigantes (GUV) con/sin encapsulación de componentes solubles en agua, utilizando electroformación24 o hinchazón asistida por gel25 y condiciones específicas para preservar la integridad de las proteínas de membrana26.
En este trabajo se presenta un protocolo para la reconstitución en LUVs de una red metabólica fuera de equilibrio que regenera ATP a través de la descomposición de L-arginina en L-ornitina27. La formación de ATP está acoplada a la producción de glicerol-3-fosfato (G3P), un componente importante para la síntesis de fosfolípidos22,28. La vía metabólica consta de dos proteínas de membrana integrales, una arginina/ornitina (ArcD) y un antiportador G3P/Pi (GlpT). Además, se requieren tres enzimas solubles (ArcA, ArcB, ArcC) para el reciclaje de ATP, y GlpK se usa para convertir el glicerol en glicerol 3-fosfato, utilizando el ATP de la descomposición de L-arginina, consulte la Figura 1 para obtener una descripción general esquemática de la vía. Este protocolo representa un buen punto de partida para la futura construcción de redes de reacción aún más complejas, para la síntesis de lípidos o proteínas o la división de células. La composición lipídica de las vesículas apoya la actividad de una amplia variedad de proteínas integrales de membrana y ha sido optimizada para el transporte de diversas moléculas dentro o fuera de las vesículas 27,29,30.
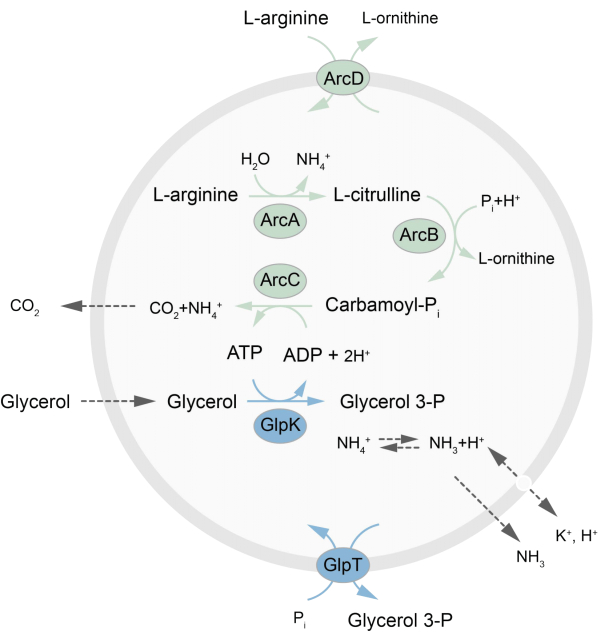
Figura 1: Descripción general de la vía para la producción de ATP y la síntesis y excreción de glicerol 3-fosfato. Haga clic aquí para ver una versión más grande de esta figura.
En resumen, se añaden proteínas de membrana purificadas (solubilizadas en dodecil-β-D-maltósido, DDM) a vesículas lipídicas preformadas que han sido desestabilizadas con Triton X-100, lo que permite la inserción de las proteínas en la membrana. Las moléculas de detergente se eliminan posteriormente (lentamente) mediante la adición de perlas de poliestireno activado, lo que da lugar a la formación de proteoliposomas bien sellados. A continuación, se pueden añadir componentes solubles a las vesículas y encapsularlos mediante ciclos de congelación y descongelación, lo que atrapa las moléculas en el proceso de fusión de la membrana. Las vesículas obtenidas son muy heterogéneas y muchas son multilaminares. Luego se extruyen a través de un filtro de policarbonato con un tamaño de poro de 400, 200 o 100 nm, lo que produce vesículas de tamaño más uniforme; Cuanto menor es el tamaño de los poros, más homogéneas y unilaminares son las vesículas, pero a costa de un menor volumen interno. Las proteínas no incorporadas y las moléculas pequeñas se eliminan de la solución externa mediante cromatografía de exclusión por tamaño. Los proteoLUV se pueden convertir en vesículas de tamaño micrométrico mediante hinchazón asistida por gel, y estos proteoGUV se recogen y atrapan en un chip microfluídico para su caracterización y manipulación microscópica. En la figura 2 se muestra una descripción general esquemática del protocolo completo.
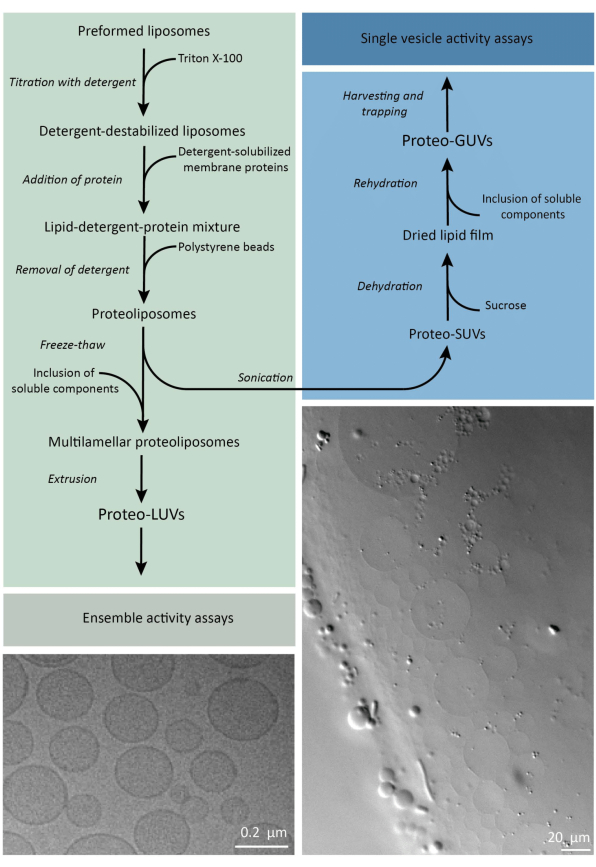
Figura 2: Resumen del protocolo para la reconstitución de proteínas de membrana y la encapsulación de enzimas y componentes solubles en agua en vesículas lipídicas de tamaño submicrométrico (LUVs) y micrométrico (GUVs). Por favor, haga clic aquí para ver una versión más grande de esta figura.
Los protocolos de reconstitución y encapsulación funcionan bien y se conserva la funcionalidad de las proteínas, pero los proteoLUVs y proteoGUVs son heterogéneos en tamaño. Los enfoques microfluídicos31,32 permiten la formación de vesículas de tamaño micrométrico que son más homogéneas en tamaño, pero la reconstitución funcional de las proteínas de membrana generalmente no es posible porque el solvente residual en la bicapa inactiva las proteínas. Los proteoLUVs varían en tamaño de 100 a 400 nm, y a bajas concentraciones de enzimas, la encapsulación puede conducir a vesículas con vías metabólicas incompletas (efectos estocásticos; ver Figura 3). Los LUV son ideales para construir módulos metabólicos específicos, como se muestra aquí para la producción de ATP y bloques de construcción como G3P. Dichos proteoLUV pueden encapsularse potencialmente en GUV y servir como compartimentos similares a orgánulos para las vesículas del huésped.
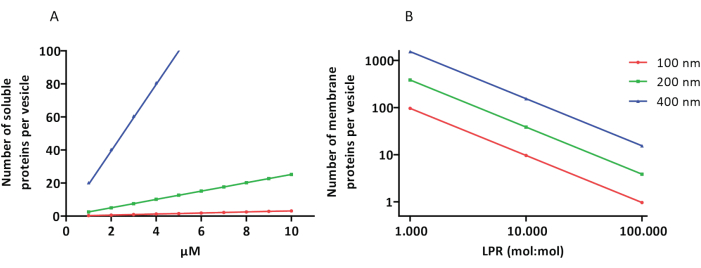
Figura 3: Número de moléculas por vesícula con un diámetro de 100, 200 o 400 nm. (A) Cuando las proteínas encapsuladas (enzimas, sondas) están en el rango de 1-10 μM. (B) La reconstitución se realiza a 1 a 1.000, 1 a 10.000 y 1 a 100.000 proteínas de membrana por lípido (mol/mol). Suponemos que las moléculas se encapsulan en las concentraciones indicadas y se incorporan a la membrana en estas proporciones proteína-lípido. En el caso de algunas enzimas, hemos visto que se unen a las membranas, lo que puede aumentar su concentración aparente en las vesículas. Abreviatura: LPR = Relación Lípido-Proteína Haga clic aquí para ver una versión más grande de esta figura.
