Protein function and activity can be altered by post translational modifications, such as phosphorylation. Many proteins can be phosphorylated by a kinase, or dephosphorylated by a phosphatase. In the case of protein phosphatase 2A (PP2A), this multisubunit phosphatase facilitates the dephosphorylation of serine and threonine residues on a broad number of phosphoproteins. Abnormal PP2A activity can lead to dysregulation of protein function which occurs in many diseases, including Parkinson's disease where aSyn can become hyperphosphorylated, and in Alzheimer's disease where tau protein can become hyperphosphorylated. This provided justification for optimizing an in-house procedure to rapidly evaluate PP2Ac activity and determine if abnormal PP2Ac activity may lead to cellular dysfunction.
Using this cell-free colorimetric assay, PP2Ac activity was measured by quantifying the amount of free PO4 cleaved from the pT substrate (Figure 1A) and compared to known picomolar amounts of PO4 in the standard curve (Figure 1B). PP2Ac activity was also measured in response to 0.0, 0.1, 0.5, 1.0, and 2.0 µM synucleins (Figure 1A), as compared to baseline PP2Ac activity in the absence of added synuclein (at the large arrow in Figure 1A).
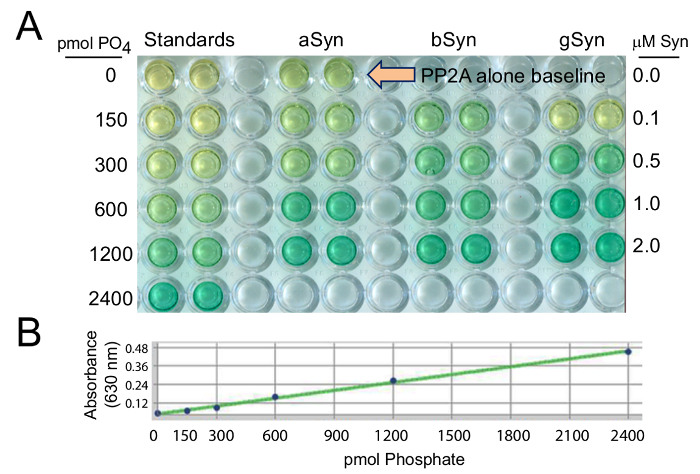
Figure 1: PP2A activation by recombinant synucleins, is measured relative to known levels of phosphate in the standards. In (A) aSyn, bSyn, and gSyn were evaluated in a range from 0 – 2 µM protein concentrations, to compare to 0 – 2400 pmol PO4 standards. (B) A standard curve for the malachite green PP2Ac assay was generated to provide known amounts of phosphate against which values obtained in experimental conditions can be calculated. In all cases, when PP2Ac activity increases, the amount of free PO4 cleaved from the pT substrate increases and produces a darker green color. Please click here to view a larger version of this figure.
The results show that addition of synucleins to PP2Ac in the initial incubation step could increase the amount of free PO4 measured, thereby demonstrating increased PP2Ac activity. The statistical analysis after many repetitions show that all three recombinant synucleins (aSyn, bSyn, gSyn) significantly increase PP2Ac activity (Figure 2). These results suggest that changes in the amount of soluble cellular synucleins can alter PP2Ac activity; and thus may impact cellular homeostasis. It is not known, however, if all three synucleins contribute to the activation of PP2Ac in vivo, though data in Figure 3 suggest that aSyn likely contributes to PP2Ac activity in vivo.
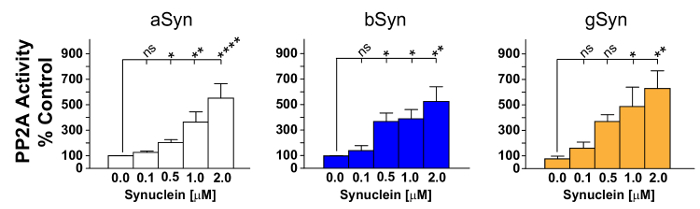
Figure 2: PP2Ac activity is stimulated by all three synucleins. Graphical representation of PP2Ac activation in response to varying concentrations of recombinant aSyn, bSyn or gSyn is shown. All three recombinant synucleins stimulated PP2Ac activity. aSyn in most experiments appeared to be slightly more potent, though ANOVAs performed using statistical software were remarkably similar overall. Data represent the mean ± SEM of 4-9 independent experiments. ns, not significant; * p <0.05; ** p <0.01; **** p <0.0001. Please click here to view a larger version of this figure.
To assess which synuclein(s) can interact with PP2Ac in brain, co-immunoprecipitation (Co-IP) assays were performed using wild type mouse brain and the PP2Ac-specific-antibody, 1D6. Proteins were then separated by SDS-PAGE and blots were probed for PP2Ac, aSyn, bSyn, and gSyn using established methods19. Recombinant synuclein controls demonstrate aSyn, bSyn and gSyn antibody specificity (Figure 3A). Co-IP data reveal that the major synuclein associated with PP2Ac in brain was aSyn, as much less bSyn came down in the co-IP (Figure 3B), and no gSyn came down in the Co-IP (data not shown). This suggests a distinctive role for endogenous aSyn in the regulation of PP2Ac in brain. It also raises the possibility that PP2Ac modulation with bSyn-based-therapies could have potential to restore PP2A activity in those with synucleinopathy, as bSyn is known to be nonamyloidogenic yet as shown here can stimulate PP2Ac30,31.
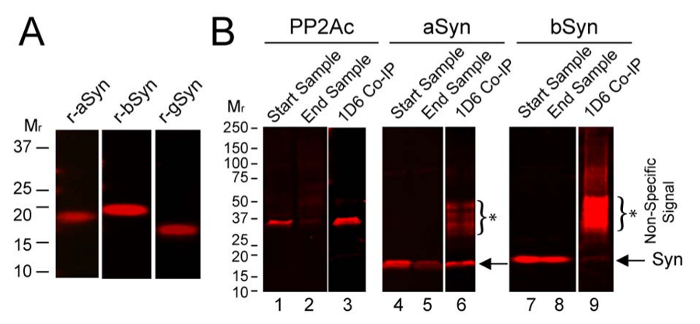
Figure 3: Interaction of PP2Ac with endogenous synucleins in mouse brain. (A) Recombinant aSyn, bSyn and gSyn were probed with synuclein specific antibodies as described below. (B) Using mouse brain homogenate Co-IP was performed with antibody 1D6, then samples were analyzed on immunoblots. Data are shown for PP2Ac, aSyn, and bSyn, but not for gSyn which did not Co-IP with PP2Ac. Start samples show relative levels of PP2Ac (lane 1), aSyn (lane 4), and bSyn (lane 7) in the initial homogenates. End samples show the remaining levels of PP2Ac (lane 2), aSyn (lane 5), and bSyn (lane 8) in homogenates after the 1D6 PP2Ac Co-IP. Significant levels of PP2Ac were immunoprecipitated using 1D6 antibody (lane 3), which also brought down ample amounts of aSyn (lane 6, at arrow) but very little bSyn (lane 9, strongly overexposed to show faint bSyn signal, at arrow). *Asterisks and brackets mark the non-specific signals on blots. See the Table of Materials for details on antibodies used. Please click here to view a larger version of this figure.
