Single cell suspensions from mechanical dissociation of testicular tissue
Figure 2 compares single cell suspensions obtained by mechanical dissociation of mouse testicular tissue under different conditions. Samples obtained by processing fresh tissue, unstained (Figure 2A) or stained with Hoechst (Figure 2B), show the presence of single cells in various stages of differentiation, and importantly, cellular structure appears to be preserved, including flagella of spermatozoa. Although some clumping and debris was observed in the stained samples, which was reduced by adding DNase after dissociation (Figure 2D), these results indicate that Hoechst staining does not significantly alter the quality of the single cell suspensions. Interestingly, single cell suspensions can also be obtained from frozen tissue for the mouse (Figure 2C) and other mammalian species – rat, dog, guinea-pig and mini-pig (Supplementary Figure 1) by mechanical dissociation. Various cell types are visible and identifiable in those samples; however, processing of frozen tissue appears to lead to increased cell death and clumping, and an overall lower cellular yield. As such, it is highly recommended to prepare single cell suspensions from fresh tissue for further downstream applications.
The quality of single cell suspensions prepared from fresh testicular tissue of mouse, rat, dog, guinea pig and mini pig, was assessed during Ho-FACS (Table 1). After exclusion of cellular debris, 95.7-98.4% of cells are singlets, and from those 86.5-93.8% are alive, as shown by quantification of PI negative cells (See Protocol). These results indicate that mechanical dissociation is a reliable method to prepare single cell suspensions for flow cytometry from testicular tissue of different mammalian species.
Ho-FACS to isolate germ cells from testicular tissue of different mammalian species
After mechanical dissociation, single cell suspensions are stained with Hoechst and PI and processed by FACS. As mentioned above, Hoechst staining allows discrimination of cells in different stages of differentiation based on chromatin quantity and structure, whereas PI staining of non-permeabilized cells separates live from dead cells. Hence, after filtering out cell debris and multicellular aggregates, the PI gate selects cells with intact membranes (PI negative; Figure 1). Live cells are then analyzed based on Hoechst fluorescence: blue is proportional to DNA content and increasing red fluorescence reflects less condensed chromatin and structural variations. As such, male germ cells of different stages are expected to cluster in specific regions of cytograms plotting the function of blue/red Hoechst fluorescence (Figure 3A).
Indeed, the Ho-FACS plots generated for the different species show the presence of distinct cell populations based on Hoechst fluorescence (Figure 3B-3C). Although these plots are sufficient to discriminate germ cell populations, a DNA content (C) gate can be defined2 to limit cross contamination. This gate is generated by a histogram of cell counts in function of Hoechst blue fluorescence intensity. For all species, three peaks can be detected and represent cells with 1C, 2C and 4C (Figure 1 and Supplementary Figures 2-5). Notably, as previously mentioned, spermatogonia are a side population and cannot be reliably gated based on histograms of DNA content. Hence, this population is gated after exclusion of dead cells, based on Hoechst blue/ red fluorescence plots (Figure 1). This strategy is represented in Figure 1 for the rat and supplementary Figures 2-5 for the remaining species. It is important to note that since Hoechst fluorescence alone can discriminate the different germ cell types, both the PI and DNA content gates are optional. They were included in this strategy to select live cells and reduce cross-contamination, respectively. As summarized in Table 1, quantification of cells in the DNA content gate reflects the relative proportion of cells in different stages of spermatogenesis. As expected, 1C cells are the most abundant and represent spermatid populations, followed by 2C cells (SPC II) and 4C cells (SPC I). Importantly, this pattern is preserved across species and small differences may reflect interspecific variability in germ cells and the cycles of the seminiferous epithelium21.
After Ho-FACS, cell integrity can be assessed using Trypan blue staining. The proportion of live cells after 1 h of FACS ranged from 27% (SCP I) to 67% (SPD) for the mouse, indicating that duration of sorting and exposure to Hoechst increases cell death. For users looking to culture the sorted cells, Hoechst concentration and duration of Ho-FACS should be adjusted to improve cell survival. Nonetheless, RNA sequencing data has been generated for single cells isolated using this method (Jung et al., unpublished), indicating that these cells are suitable for molecular studies of germ cell biology. This dataset was used to detect potential contamination of germ cell populations with somatic cells (Supplementary Figure 6), which was estimated to be less than 5%.
Morphologic evaluation of sorted germ cell populations
As germ cells undergo drastic morphological changes throughout spermatogenesis, it is possible to estimate the purity of isolated populations by microscopy. As a reference, Figure 4 illustrates the general morphology of different male germ cells types from 4 mammalian species (from Lima, et al. 5). Chromatin distribution and condensation, as well as cell size and shape, show unique and well characterized patterns in different cell types. SPG have distinct pericentric heterochromatin, which stains brightly for Hoechst, and are small and round in shape (Spg in Figure 4B). Spermatocytes are larger granulated cells. Nuclei of SPC I are easily identifiable, with chromatin variations characteristic of meiotic cells (Spc I in Figure 4B), whereas SPC II are binucleated or in diakinesis (Spc II in Figure 4B). Finally, SPD are small haploid cells with round or elongated shape (Spd in Figure 4B). Notably, rSPD are similar to SPG in size and shape, but clearly distinguished by the presence of localized chromocenters. Based on these characteristics, sorted cell populations were evaluated for the enrichment of specific cell types (Table 1). SPD and SPC II populations were highly enriched for the cell type of interest, whereas SPC I and SPG showed some degree of contamination with other cell types, especially for the Guinea Pig and Mini Pig samples. Interestingly, for the dog sample the gating strategy was sufficient to discriminate round from elongating spermatids (Figure 3C; Supplementary Figure 3; Table 1). However, this was not the case for most species, where spermatid subpopulations appear to overlap.
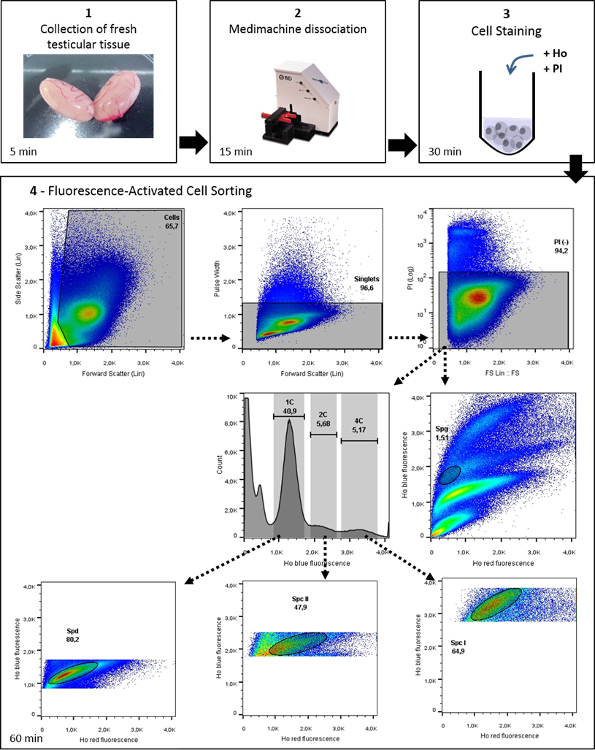
Figure 1: Workflow of Ho-FACS isolation of mammalian male germ cells. This image illustrates the general protocol for germ cell isolation of mammalian germ cells, with representative data from the application of this protocol to rat testis. Ho: Hoechst. From Lima, et al. 5 with permission. Please click here to view a larger version of this figure.
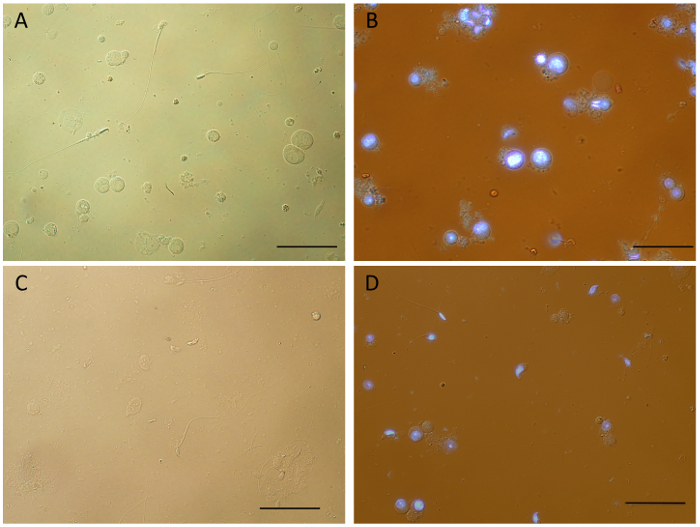
Figure 2: Single cell suspensions generated by mechanical dissociation of mouse testicular tissue. Samples obtained from fresh tissue (A-B) show a variety of intact germ cell types and very little debris. When compared to unstained cells (A), some cell clumping was observed for Hoechst stained cell suspensions (B), which can be reduced by adding DNase (final concentration of 10 µg/mL) to the sample immediately after dissociation (D). Frozen testicular tissue can also be used to obtain single cell suspensions by mechanical dissociation (C). However, it appears that the process of flash freezing and thawing prior to tissue dissociation results in more debris, cell death and overall less cellular yield. After dissociation (See Protocol), samples were spun for 10 min at 4 °C and re-suspended in 1x DMEM before mounting the slides. Black bars = 50 µm. Images obtained using an upright white light microscope equipped with a UV lamp. Please click here to view a larger version of this figure.
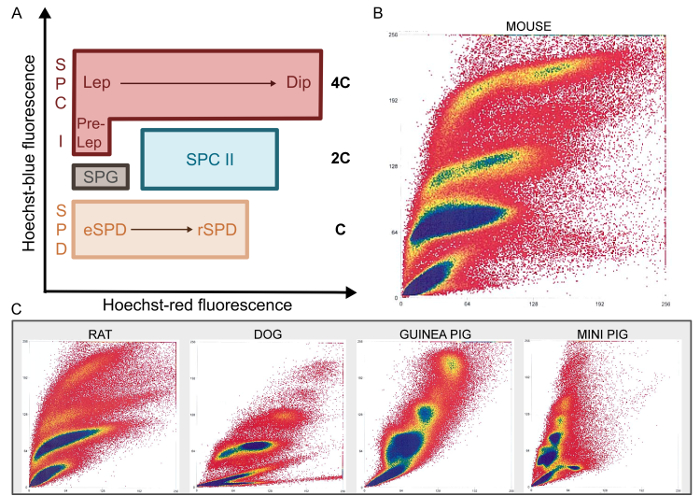
Figure 3: Ho-FACS of mammalian germ cells. Specific populations present unique patterns of Hoechst fluorescence during FACS (A). As Hoechst blue fluorescence reflects variations in chromatin content clusters of haploid (C); diploid (2C) and tetraploid (4C) cells are located in areas of the FACS plot with increasing Hoechst blue fluorescence. Plots generated as a function of Hoechst blue/red fluorescence show clusters of distinct cell populations for the different mammalian species tested (B-C). SPG: Spermatogonia; SPC I: Primary spermatocytes; Pre-Lep: Pre-Leptotene spermatocytes; Lep: Leptotene spermatocytes; Dip: Diplotene spermatocytes; SPC II: Secondary spermatocytes; SPD: Spermatids; eSPD: elongating spermatids; rSPD: round spermatids. Adapted from Lima, et al. 5 with permission. Please click here to view a larger version of this figure.
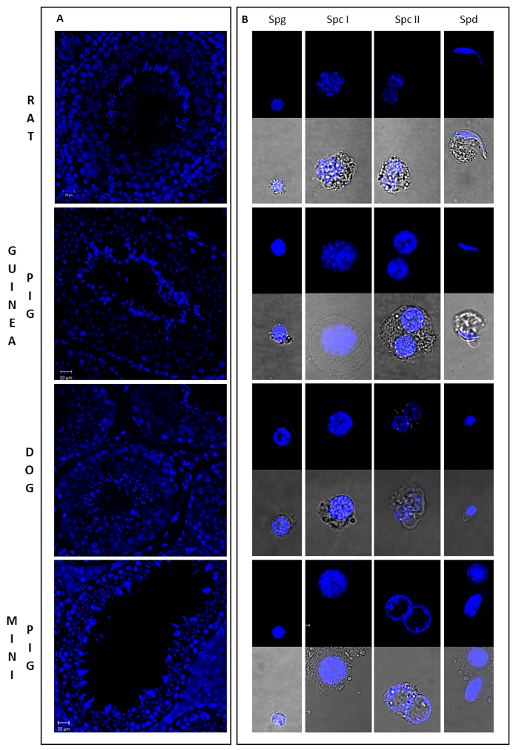
Figure 4: Morphology of germ cells in different developmental stages from four mammalian species. This figure shows sections of testicular tissue stained with Hoechst (A) and the general aspect of different male germ cell types sorted by Ho-FACS (B) from rat, guinea pig, dog and mini pig. As cell size, shape and chromatin conformation varies dramatically throughout spermatogenesis, these can be used to assign cells to different stages of differentiation. Spermatogonia (Spg) are small and round cells with distinct pericentric heterochromatin and exhibit a high intensity of Hoechst fluorescence. Spermatocytes are the largest germ cells and show variable chromatin conformations. Nuclei of primary spermatocytes (Spc I) exhibit chromatin variations characteristic of meiotic cells, whereas secondary spermatocytes (Spc II) are binucleated or in diakinesis. Spermatids (Spd) have round or elongated shape, depending on the stage of spermiogenesis and are small haploid cells. Although round spermatids and spermatogonia are similar in size and shape, the former can be distinguished by the presence of localized chromocenters. Slides were prepared for each sorted cell type after Ho-FACS and were visualized in a confocal microscope: 63X magnification lens, with (lower panel) or without (upper panel) white light transmission.From Lima, et al. 5 with permission. Please click here to view a larger version of this figure.
| % cells in DNA content gate | Purity of sorted germ cell populations (%) | |||||||||||
| Species | % Singlets | % Live cells | 1C | 2C | 4C | Spg | Spc I | Spc II | Spd | rSpd | eSpd | |
| Mouse | 98.4 | 92.5 | 34.7 | 3.9 | 3.72 | 74 | 82 | 87.5 | 95.2 | 95* | 92* | |
| Rat | 95.7 | 93.8 | 37.7 | 5.3 | 5.4 | 83 | 81 | 82 | 87 | . | . | |
| Guinea Pig | 96.2 | 92.1 | 39.3 | 7.6 | 5.8 | 48 | 68.7 | 85 | 87 | . | . | |
| Dog | 97.9 | 86.5 | 16.4 | 3 | 0.5 | 78 | . | 87 | . | 91 | 81 | |
| Mini Pig | 95.9 | 93.2 | 26.9 | 6.4 | 3.5 | 49 | 52 | 82 | 92 | . | . | |
| * Obtained by enzymatic dissociation and gated based on FSC&SSC parameters (from Lima et al. 2016, with permission) | ||||||||||||
Table 1: Statistics of Ho-FACS of male germ cell suspensions obtained by mechanical dissociation.
Supplementary Figure 1: Single cell suspensions obtained by mechanical dissociation of frozen testicular tissue. Mechanical dissociation allows the generation of single cell suspensions from testicular tissue of different mammalian species without the need of species-specific protocols. Different panels show the cell suspensions obtained for 4 mammalian species. Samples were obtained by mechanical dissociation of frozen testicular tissue and stained with Hoechst. SPG: Spermatogonia; SPC I: Primary Spermatocyte; SPC II: Secondary Spermatocyte; SPD: Spermatid; SPZ: Spermatozoa. Slides were visualized in a confocal microscope with (lower panel) or without (upper panel) white light transmission. Scale bars =20 µm. Please click here to download this file.
Supplementary Figure 2: Gating strategy for separation of live male germ cells by Ho-FACS of mouse testicular single cell suspensions. Live cells (PI negative) are sorted according to Hoechst fluorescence. Spermatogonia (SPG) are identified by plotting directly Hoechst-blue and red fluorescence intensities. A DNA content gate is used to sort cells based on Hoechst blue fluorescence: haploid (C); diploid (2C) and tetraploid (4C). Cells in each peak of the histogram are then sorted by plotting the function of Hoechst blue and red fluorescence. SPG: Spermatogonia; SPD: Spermatids; SPC II: Secondary spermatocytes; SPC I: Primary spermatocytes. Please click here to download this file.
Supplementary Figure 3: Gating strategy for separation of live male germ cells by Ho-FACS of dog testicular single cell suspensions. Live cells (PI negative) are sorted according to Hoechst fluorescence. Spermatogonia (SPG) are identified by plotting directly Hoechst-blue and red fluorescence intensities. A DNA content gate is used to sort cells based on Hoechst blue fluorescence: haploid (C); diploid (2C) and tetraploid (4C). Cells in each peak of the histogram are then sorted by plotting the function of Hoechst blue and red fluorescence. The peak containing haploid cells can be subdivided according to the range of Hoechst blue intensity and represent two subpopulations of spermatids: elongating spermatids (gate C') and round spermatids (gate C''). SPG: Spermatogonia; eSPD: elongating spermatids; rSPD: round spermatids; SPC II: Secondary spermatocytes; SPC I: Primary spermatocytes. Please click here to download this file.
Supplementary Figure 4: Gating strategy for separation of live male germ cells by Ho-FACS of guinea pig testicular single cell suspensions. Live cells (PI negative) are sorted according to Hoechst fluorescence. Spermatogonia (SPG) are identified by plotting directly Hoechst-blue and red fluorescence intensities. A DNA content gate is used to sort cells based on Hoechst blue fluorescence: haploid (C); diploid (2C) and tetraploid (4C). Cells in each peak of the histogram are then sorted by plotting the function of Hoechst blue and red fluorescence. SPG: Spermatogonia; SPD: Spermatids; SPC II: Secondary spermatocytes; SPC I: Primary spermatocytes. Please click here to download this file.
Supplementary Figure 5: Gating strategy for separation of live male germ cells by Ho-FACS of mini pig testicular single cell suspensions. Live cells (PI negative) are sorted according to Hoechst fluorescence. Spermatogonia (SPG) are identified by plotting directly Hoechst-blue and red fluorescence intensities. A DNA content gate is used to sort cells based on Hoechst blue fluorescence: haploid (C); diploid (2C) and tetraploid (4C). Cells in each peak of the histogram are then sorted by plotting the function of Hoechst blue and red fluorescence. SPG: Spermatogonia; SPD: Spermatids; SPC II: Secondary spermatocytes; SPC I: Primary spermatocytes. Please click here to download this file.
Supplementary Figure 6: Examination of somatic cell contamination in Ho-FACS sorted populations. t-distributed stochastic neighbor embedding (t-SNE) plot of approximately 600 single-cell transcriptome from three FACS subpopulation gates (A-B). On the t-SNE plot, each point represents the unique position of a single cell in a transcriptome space and each cell is labeled with its FACS subpopulation gate origin (A). Visual quantification of cell-type specific markers on the FACS single-cell t-SNE plot (B). Contamination from somatic cells is estimated to be less than 5% and is not specific to one Ho-FACS population gate. Please click here to download this file.
