Figure 1 represents a schematic of the described steps above. Isolated murine spleens are sorted for CD11c+ DCs by magnetic cell sorting and plated in either normal salt media (NS; 150 mmol NaCl) or high salt media (HS; 190 mmol NaCl) for 48 h. CD11c+ DCs are then adoptively transferred by retro-orbital injection to naïve recipient mice. Ten days later, mice are implanted with osmotic minipumps for low-dose angiotensin II (140 ng/kg/min) infusion for 14 days. During the 14 day infusion of angiotensin II, blood pressure is recorded by radio telemetry or tail-cuff plethysmography.
A typical blood pressure analysis is represented in Figure 2 (modified from Barbaro et al.9) following adoptive transfer of DCs and osmotic minipumps for low-dose angiotensin II infusion are implanted. Of note, this low dose of angiotensin II (140 ng/kg/min) is a sub pressor dose that does not increase blood pressure in a normal mouse. Mice receiving normal salt treated CD11c+ DCs (black circles) maintain a normal blood pressure during the angiotensin II infusion, while mice receiving high salt treated CD11c+ DCs (red circles) exhibit an increase in systolic blood pressure. Figure 2 illustrates that high salt treated CD11c+ DCs prime hypertension in response to a sub pressor dose of angiotensin II.
To determine the purity of the isolated CD11c+ cells, we performed flow cytometry analysis using a gating strategy illustrated in Figure 3A. We found that compared to the total splenocytes, we achieved increased enrichment of CD11c+ cells (Figure 3B). We utilized different CD11c microbead concentrations to troubleshoot the manufacturer's protocol in Figure 4. Following magnetic separation of splenocytes using 100 μL (Figure 4A), 200 μL (Figure 4B), or 300 μL (Figure 4C) of CD11c microbeads yields about 65%, 55%, and 50% of CD11c+ positive cells respectively. We then included an FcR blocker (5 μL/spleen) + 100 μL of CD11c microbead, magnetically separated, and found that blockade of FcR yields approximately 65% CD11c positive cells (Figure 4D). In additional experiments, we incubated splenocytes in 100 μL CD11c microbeads and magnetic separated through an LS column. We then incubated the separated cells with an additional 100 μL of CD11c microbeads and again separated through an LS column. Double isolation of splenocytes yielded 92% CD11c+ cells (Figure 4E).
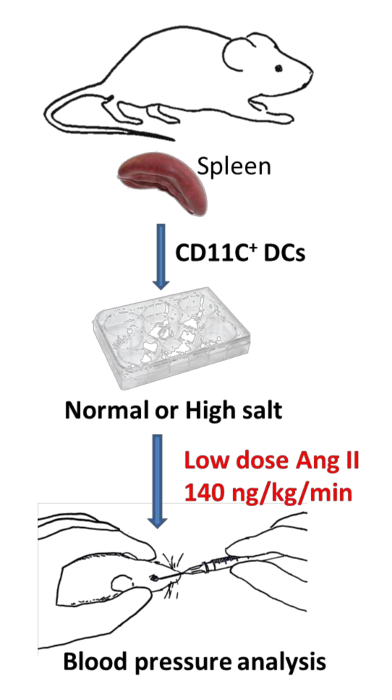
Figure 1: Illustration of adoptive transfer of in vitro treated CD11c+ DCs. CD11c+ DCs are isolated from spleens of mice and plated in either normal salt or high salt media for 24 h. CD11c+ DCs are then adoptively transferred retro-orbitally into naïve recipient mice. An osmotic minipump infusing low-dose angiotensin II is implanted and blood pressure is monitored for 14 days. This figure is adapted from Barbaro et al.)9. Please click here to view a larger version of this figure.
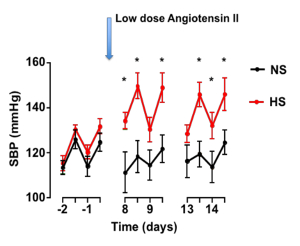
Figure 2: The effect of high salt treated CD11c+ DCs on systolic blood pressure. Dendritic cells were isolated and cultured in normal salt (NS) or high salt (HS) for 48 h. DCs were adoptively transferred to wild type naïve mice. Ang II minipumps (140 ng/min) were implanted and blood pressure monitored by radio telemetry. Systolic blood pressure (SBP) measured by radio telemetry (mean ± SEM). This figure is adapted from Barbaro et al.9 Please click here to view a larger version of this figure.
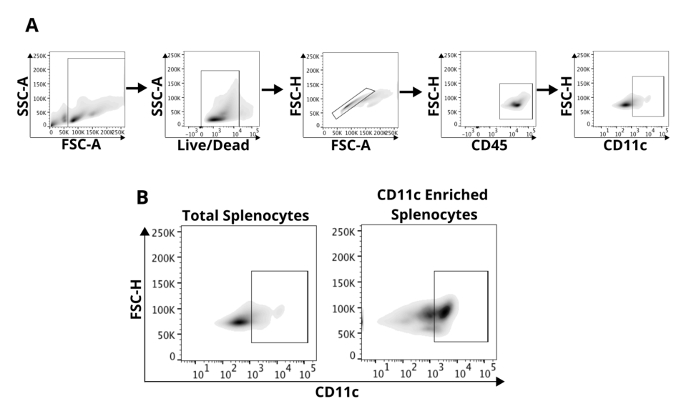
Figure 3: Flow cytometric analysis of magnetically separated splenocytes. (A) Gating strategy defining analysis of CD11c+ DC population. Cells are separated from debris and living cells are selected based upon exclusion of Live/Dead cell stain. Singlet cells are selected and then analyzed for CD45 positivity. CD45 cells are then analyzed for CD11c. (B) Following magnetic separation, there is significant enrichment of CD11c+ Cells. Please click here to view a larger version of this figure.
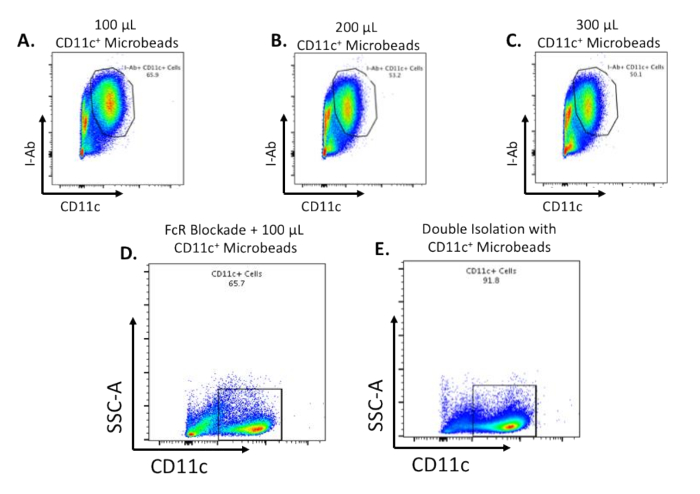
Figure 4: Flow cytometric analysis of magnetically separated CD11c+ splenocytes after different isolation protocols. Cells were separated from debris and living cells. Live cells were analyzed for I-Ab and CD11c positivity. (A) % of CD11c+ cells following magnetic separation that were isolated with 100 μL, (B) 200 µL, (C) 300 μL of CD11c microbeads. (D) % of CD11c+ cells following magnetic separation that were isolated with blockade of FcR (anti-FcR; 5 μL) + 100 μL CD11c microbeads. (E) % of CD11c+ cells following magnetic separation that were isolated with double magnetic cell sorting. Please click here to view a larger version of this figure.
