The production of mucoid EPS could be observed in B. longum cultures on PYG plates after anaerobic incubation for 72 h (Figure 1A). Centrifugation of culture scrapes, followed by ethanol precipitation and drying, resulted in the collection of cellulose-like EPS (Figure 1B). Dried EPS and soluble starch were then used as carbon sources for fermentation cultures. TLC was used for oligosaccharide separation and purity analysis due to its low cost and rapid results turnaround18. Although the degradation rate of starch by human fecal microbiota was faster than that of Bif EPS (Figure 2), Bif EPS degradation was clearly observed for some EPS-inoculated samples.
Community compositional analysis via 16S rRNA gene high-throughput sequencing and principal coordinate analysis (PCoA) was then performed to investigate the effects of Bif EPS on human gut microbiota. Samples from the VI_Bif and VI_Starch groups clustered separately from each other in the PCoA analysis (Figure 3A), indicating that EPS and starch availability differentially shape human fecal bacterial communities. Linear discriminant analysis effect size (LEfSe) was further used to distinguish the specific bacterial taxa that differed between the VI_Bif and VI_Starch treatments. The genera Collinsella, Coprococcus, Parabacteroides, and Rhodopseudomonas were significantly more abundant in the VI_Bif samples than in the VI_Starch samples (Figure 3B). Furthermore, GC measurements were made for several SCFAs to evaluate their production following the addition of different carbon sources. SCFAs that were measured from fermentation cultures included acetic, propionic, isobutyric, butyric, isovaleric, and valeric acids. Following fermentation for 24 h and 48 h, five of the six aforementioned SCFA concentrations were similar among treatments and not statistically different between the VI_Bif, VI_Starch, and VI groups. However, propionic acid concentrations were significantly higher in the VI_Bif group than in the VI_Starch group (Figure 4).

Figure 1: EPS produced by B. longum.
Frozen B. longum was restored in Bifidobacterium medium broth and then streaked onto PYG plates, followed by anaerobic incubation at 37 °C for 72 h (A). The EPS produced by bacterial cultures were scraped from plate cultures, precipitated using ethanol, and dried overnight using a speed vacuum (B). Please click here to view a larger version of this figure.

Figure 2: TLC analysis of in vitro EPS and starch degradation by human gut microbiota.
TLC analysis was conducted on 0.2 μL samples collected at 24 h and 48 h from each fermentation culture grown under anaerobic conditions. VI, VI_Starch, and VI_Bif indicate VI media, VI media + starch supplement, and VI media + EPS supplement, respectively. The numbers 1–12 indicate fecal bacterial samples from the 12 volunteers that were used to inoculate the fermentation experiments. The control group represents treatment without additional carbon supplements. This figure is modified from Yin et al.11. Please click here to view a larger version of this figure.
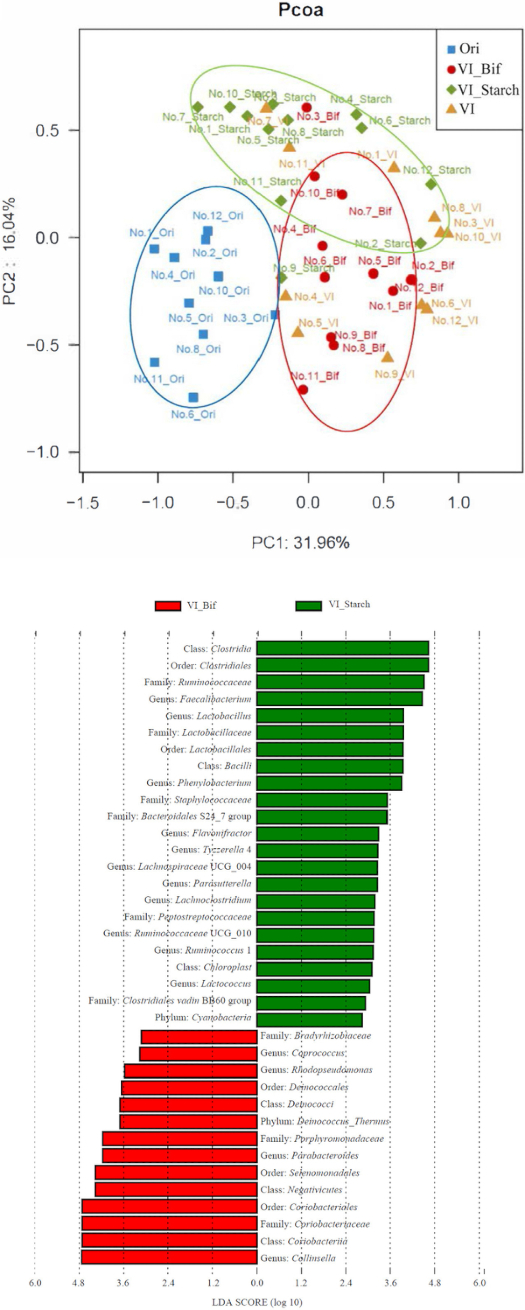
Figure 3: Effects of Bif EPS availability on human gut microbiota communities.
(A) PCoA plot of gut microbiota community compositional dissimilarities based on the unweighted UniFrac metric. (B) LEfSE analysis of bacterial taxa that were differentially abundant among treatment groups. A cutoff of p < 0.05 was used to assess the statistical significance of bacterial taxonomic differences among groups. Ori indicates the gut microbiota of the volunteer fecal samples. VI_Bif and VI_Starch indicate the gut microbiota from fermentation samples using VI media with EPS and starch as carbon substrates, respectively. VI represents the control group with gut microbiota inoculated fermentations in VI media without supplementation of other carbohydrates. This figure is modified from Yin et al.11 Please click here to view a larger version of this figure.
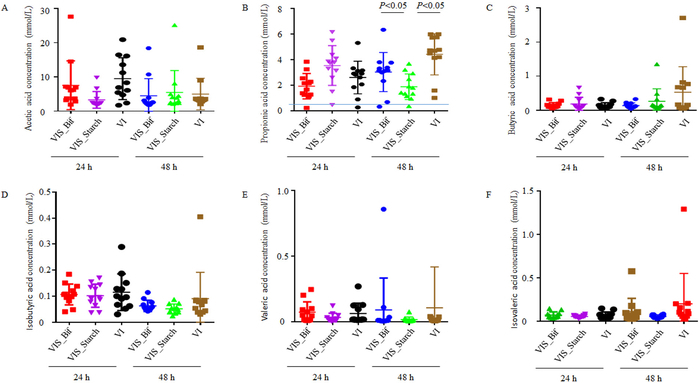
Figure 4: Effects of EPS availability on SCFA production after 24 h and 48 h of fermentation.
Acetic, propionic, isobutyric, butyric, isovaleric, and valeric acids were detected using gas chromatography. VI_Bif, VI_Starch, and VI indicate the samples that were collected after cultivation using VI media + EPS, VI media + starch, and VI media, respectively. All samples were measured in triplicate. The figures were generated using GraphPad Prism Version 5.01. Panels represent organic acid concentrations within each fermented sample for A, acetic acid; B, propionic acid; C, isobutyric acid; D, butyric acid; E, isovaleric acid; and F, valeric acid. This figure is modified from Yin et al.11 Please click here to view a larger version of this figure.
