Detecting Virus and Salivary Proteins of a Leafhopper Vector in the Plant Host
Summary
This protocol demonstrates how to use the plant host to detect salivary proteins of leafhopper and plant viral proteins released by leafhopper vectors.
Abstract
Insect vectors horizontally transmit many plant viruses of agricultural importance. More than one-half of plant viruses are transmitted by hemipteran insects that have piercing-sucking mouthparts. During viral transmission, the insect saliva bridges the virus-vector-host because the saliva vectors viruses, and the insect proteins, trigger or suppress the immune response of plants from insects into plant hosts. The identification and functional analyses of salivary proteins are becoming a new area of focus in the research field of arbovirus-host interactions. This protocol provides a system to detect proteins in the saliva of leafhoppers using the plant host. The leafhopper vector Nephotettix cincticeps infected with rice dwarf virus (RDV) serves as an example. The vitellogenin and major outer capsid protein P8 of RDV vectored by the saliva of N. cincticeps can be detected simultaneously in the rice plant that N. cincticeps feeds on. This method is applicable for testing the salivary proteins that are transiently retained in the plant host after insect feeding. It is believed that this system of detection will benefit the study of hemipteran-virus-plant or hemipteran-plant interactions.
Introduction
The vector-host transmission mode of arboviruses, a fundamental problem, is at the frontier of biological science. Many plant viruses of agricultural importance are horizontally transmitted by insect vectors1. More than one-half of plant viruses are vectored by hemipteran insects, including aphids, whiteflies, leafhoppers, planthoppers, and thrips. These insects have distinct features that enable them to efficiently transmit plant viruses1. They possess piercing-sucking mouthparts and feed on the sap from phloem and xylem, and secrete their saliva1,2,3,4. With the development and improvement of techniques, the identification and functional analyses of salivary components are becoming a new focus of intensive research. The known salivary proteins in saliva include numerous enzymes, such as pectinesterase, cellulase, peroxidase, alkaline phosphatase, polyphenol oxidase, and sucrase, among others5,6,7,8,9,10,11,12,13. The proteins in saliva also include elicitors that trigger the host defense response, thereby altering the performance of insects, and effectors that suppress the host defense, which enhances insect fitness and components that induce host pathological responses14,15,16,17. Therefore, saliva proteins are vital materials for communication between insects and hosts. During the transmission of viruses, the saliva secreted by the salivary glands of piercing-sucking viruliferous insects also contains viral proteins. Viral components utilize the flow of saliva to release them from the insect to the plant host. Therefore, the insect saliva bridges the virus-vector-host tritrophic interaction. Investigating the biological function of saliva proteins secreted by viruliferous insects helps to understand the relationship of virus-vector-host.
For animal viruses, it is reported that the saliva of mosquitoes mediates the transmission and pathogenicity of West Nile virus (WNV) and Dengue virus (DENV). The saliva protein AaSG34 promotes dengue-2 virus replication and transmission, while the saliva protein AaVA-1 promotes DENV and Zika virus (ZIKV) transmission by activating autophagy18,19. The saliva protein D7 of mosquitoes can inhibit DENV infection in vitro and in vivo via direct interaction with the DENV virions and recombinant DENV envelope protein20. In plant viruses, the begomovirus tomato yellow leaf curl virus (TYLCV) induces the whitefly salivary protein Bsp9, which suppresses the WRKY33-mediated immunity of plant host, to increase the preference and performance of whiteflies, eventually increasing the transmission of viruses21. Because studies of the role that insect salivary proteins play in plant hosts have lagged behind those of animal hosts, a stable and reliable system to detect the salivary proteins in plant hosts is urgently required.
The plant virus known as rice dwarf virus (RDV) is transmitted by the leafhopper Nephotettix cincticeps (Hemiptera: Cicadellidae) with high efficiency and in a persistently propagative manner22,23. RDV was first reported to be transmitted by an insect vector and causes a severe disease of rice in Asia24,25. The virion is icosahedral and double-layered spherical, and the outer layer contains the P8 outer capsid protein22. The circulative transmission period of RDV in N. cincticeps is 14 days26,27,28,29,30. When the RDV arrives at salivary glands, virions are released into saliva-stored cavities in the salivary glands via an exocytosis-like mechanism23. The vitellogenin (Vg) is the yolk protein precursor essential for oocyte development in female insects31,32,33. Most insect species have at least one Vg transcript of 6-7 kb, which encodes a precursor protein of approximately 220 kDa. The protein precursors of Vg can usually be cleaved into large (140 to 190 kDa) and small (<50 kDa) fragments before entering the ovary18,19. Previous proteomic analysis revealed the presence of the peptides derived from Vg in the secreted saliva of the leafhopper Recilia dorsalis, although their function is unknown (unpublished data). It is newly reported that Vg, which is orally secreted from planthoppers, functions as an effector to damage the defenses of plants34. It is unknown whether the Vg of N. cincticeps could also be released to the plant host with salivary flow, and then could play a role in the plant to interfere with plant defenses. To address whether N. cincticeps exploits salivary proteins, such as Vg, to inhibit or activate plant defenses, the first step is identifying proteins released to the plant during feeding. Understanding the method to identify the salivary proteins present in the plant is potentially essential to explain the function of saliva proteins and the interactions between Hemiptera and plants.
In the protocol presented here, N. cincticeps is used as an example to provide a method to examine the presence of salivary proteins in the plant host introduced through insect feeding. The protocol primarily details the collection and detection of salivary proteins and is helpful for further investigation on most hemipterans.
Protocol
The non-viruliferous adult leafhoppers were propagated in the Vector-borne Virus Research Center in Fujian Agriculture and Forestry University, China.
1. Nonviruliferous insect rearing
- Rear the adults on rice seedlings in a cube cage that is 40 cm x 35 cm x 20 cm (length x width x height). Keep one side of the cage covered with an insect-proof net for ventilation.
- Keep the cages with leafhoppers in an incubator that contains an in-built humidity controller at 26 °C with a relative humidity of 60-75% under a photoperiod of 16 h light and 8 h dark.
- Use an aspirator to gently transfer all the adults from their cage into a new cage that contains fresh rice seedlings each week.
- Let more than 200 adults mate and lay eggs in the rice.
- Retain the old rice seedlings for the nymphs to emerge. Rear these new nonviruliferous nymphs to the 2-instar stage.
2. Virus acquisition and the collection of viruliferous insects
- Carefully transfer the 2-instar nonviruliferous nymphs to a glass culture tube (2.5 cm in diameter by 15 cm high) for 1-2 h for starvation using the aspirator.
- Release the nymphs to a cage that contains an RDV-infected rice plant grown in a pot.
- Allow the nymphs to feed on the infected rice plant for 2 days.
NOTE: Carefully water the rice plant and avoid washing away the nymphs. The 2-instar nymphs are approximately 1.6-2 mm long.
- Carefully transfer these nymphs to a new cage that contains fresh virus-free rice seedlings with a relative humidity of 60-75% under a photoperiod of 16 h light and 8 h dark. Allow the nymphs to feed on the infected rice plant for 12 days to complete the circulative transmission period of RDV.
3. Collection of salivary proteins using a feeding cage
- Prepare five small pipe-like feeding cages (2.5 cm in diameter by 4 cm high) in which one end is covered with insect-proof netting.
- Confine 15-20 leafhoppers in each feeding cage, and then cover the other end of the cage with a thin foam mat.
- Fix one rice seedling (5-6 cm high) between the end of cage and a foam mat with tapes. Ensure that the leafhoppers in the feeding cage can feed on the rice seedlings exposed to the interior of the cage.
- Immerse the seedling roots in water so that the rice plant will remain alive. Allow the leafhoppers to feed on them for 2 days.
- Remove the leafhoppers from their feeding cages and collect the rice seedlings on which they fed. Cut the parts of seedlings outside of the cage and recover the feeding regions of seedlings.
NOTE: This sample can be stored at -80 °C for 3 months at the most, if it is not instantly used for detection.
4. Reagent preparation
- Dissolve 15.1 g of Tris-base, 94 g of glycine, and 5 g of SDS in 1 L of sterile water to prepare 5xTris-glycine buffer. Dilute 200 mL of 5xTris-glycine buffer with 800 mL of sterile water to prepare 1xTris-glycine buffer (see Table 1 for buffer composition).
- Dissolve 80 g of NaCl, 30 g of Tris-base, and 2 g of KCl in 1 L of sterile water to prepare 10xTris-buffered saline (TBS) buffer. Autoclave the solution at 121 °C for 15 min.
- Dissolve 8 g of SDS, 4 mL of ß-mercaptoethanol, 0.02 g of bromophenol blue, and 40 mL of glycerol in 40 mL of 0.1 M Tris-HCl (pH 6.8) to prepare 4x protein sample buffer.
- Mix 800 mL Tris-glycine buffer with 200 mL methanol to prepare the transfer buffer.
- Add 100 mL 10xTBS solution and 3 mL Tween 20 to 900 mL sterile water to prepare the TBS buffer with Tween 20 (TBST) solution.
5. Western blotting to detect the saliva and viral proteins
- Grind 0.1 g of the rice samples with liquid nitrogen until the tissue becomes a powder. Add 200 µL of 4x protein sample buffer to the sample and boil it for 10 min. Centrifuge the samples at 12,000 x g for 10 min at room temperature.
- Remove the supernatant and place it in a new vial. Load 10 µL of the sample into an SDS-PAGE gel, and run it in Tris-glycine buffer at 150 V for 45-60 min.
NOTE: The residue after centrifugation can be discarded.
- Remove the supernatant and place it in a new vial. Load 10 µL of the sample into an SDS-PAGE gel, and run it in Tris-glycine buffer at 150 V for 45-60 min.
- Put a 0.45 µm nitrocellulose membrane and other sandwich supplies in the Transfer buffer for 30 min.
NOTE: This step can be done before the gel has completed its run. - Sandwich the gel and transfer it for 90-120 min at 100 V in the Transfer buffer.
- Take the membrane and place it in 7% non-fat dry milk blocking solution in TBST solution for 20 min. Add the specific antibody against RDV P8 or Vg to a 7% solution of non-fat dry milk in TBST. Incubate with the antibody for staining the membrane for 2 h or overnight.
- Wash the membrane with TBST solution three times, with 5 min washing each time.
- Add the goat anti-rabbit IgG as a secondary antibody to 7% non-fat dry milk with TBST. Incubate with the antibody for 60-90 min at room temperature.
- Wash the membrane with TBST solution three times for 5 min each time.
- Use the ECL Western kit for the chemiluminescent method. Mix Detection Reagents 1 and 2 in the kit at a ratio of 1:1 in a tube. Put the mixed reagent onto the membrane and incubate the blot for 5 min.
- Drain the excess reagent and take a colorimetric picture of the chemiluminescent picture. Combine them to see the ladder with protein bands.
Representative Results
Figure 1 illustrates all of the steps in this protocol: insect rearing, virus acquisition, the collection of salivary proteins via rice feeding, and the western blot. The western blots results showed that specific and expected bands of approximately 220 kDa were observed in the samples of feeding rice and salivary glands of insects on the membrane incubated with antibodies against Vg. In contrast, no band was observed in the non-feeding rice sample. The result in Figure 2 indicates that Vg was released to the plant host as a salivary protein. On the membrane incubated with antibodies against RDV P8, a specific and expected band of approximately 46 kDa was also observed in the samples from rice that had been subjected to feeding and the bodies of viruliferous insect bodies. In contrast, no band was observed in the non-feeding rice sample and the nonviruliferous leafhopper bodies, as shown in Figure 3. This result proved that the viral proteins could also be detected in the feeding plants.
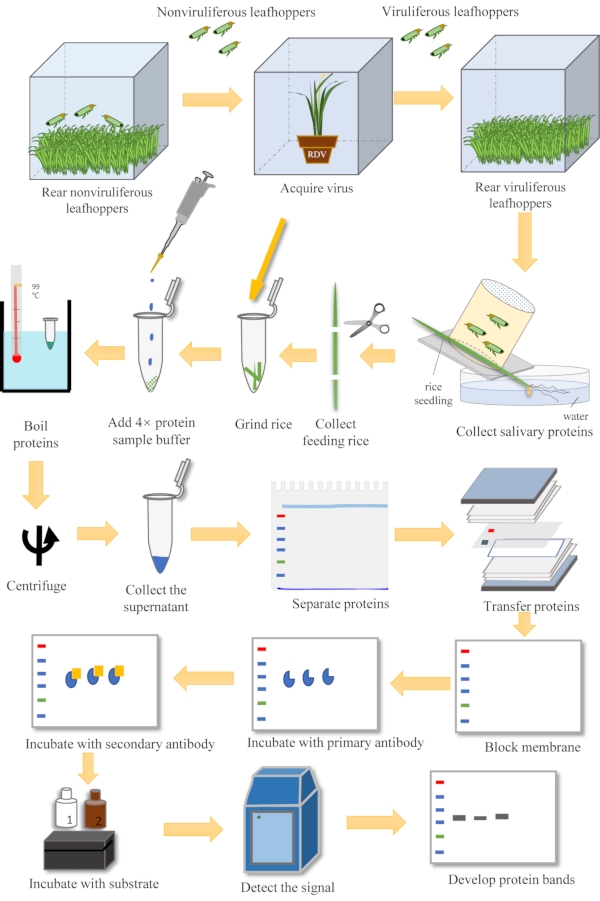
Figure 1: Overview of the steps in the detection of salivary proteins. The steps include insect rearing, virus acquisition, salivary proteins collection via plant feeding, and western blotting. Please click here to view a larger version of this figure.
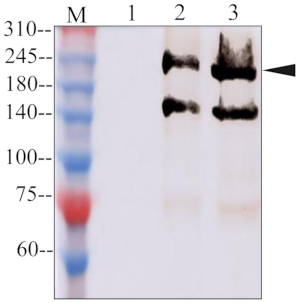
Figure 2: Western blot assay of Vg in plant and insect samples. Lane 1, non-feeding plants; Lane 2, viruliferous leafhopper-feeding plants; Lane 3, viruliferous leafhopper salivary glands; Lane M, marker. Please click here to view a larger version of this figure.

Figure 3: Western blot assay of P8 in plant and insect samples. Lane 1, non-feeding plants; Lane 2, viruliferous leafhopper-feeding plants; Lane 3, viruliferous leafhopper bodies; Lane 4, nonviruliferous leafhopper bodies; M, marker. Please click here to view a larger version of this figure.
| Buffer | Composition | Comments/Description |
| 5x Tris-glycine buffer | 15.1 g Tris base 94 g glycine 5 g SDS in 1 L sterile water |
Stock solution |
| 1x Tris-glycine buffer | 200 mL of 5x Tris-glycine buffer 800 mL sterile water |
Work solution, for SDS-PAGE |
| 10x Tris-buffered saline (TBS) buffer | 80 g NaCl 30 g Tris base 2 g KCl in 1 L sterile water |
Stock solution |
| TBS with Tween 20 (TBST) solution | 100 mL 10x TBS solution 3 mL Tween 20 900 mL sterile water |
Work solution |
| 4x protein sample buffer | 8 g SDS 4 mL β-mercaptoethanol 0.02 g bromophenol blue 40 mL glycerol in 40 mL 0.1 M Tris-HCl (pH 6.8) |
For protein extraction |
| Transfer buffer | 800 mL Tris-glycine buffer 200 mL methanol |
For protein transfer |
Table 1: Buffers, solutions, and reagents used in the study. The composition of the buffers and the solutions, along with their usage, are listed.
Discussion
The saliva directly secreted by the salivary glands of the piercing-sucking insects plays a pivotal role because it predigests and detoxifies the host tissues and vectors' cross-kingdom biological factors into the hosts1,3,4. The cross-kingdom biological factors, including elicitors, effectors, and small RNA, are critical for insect-host communication14,15,16. Therefore, uncovering more varieties and functions of salivary components will promote understanding the relationship between insects and hosts. Here, a system of detection for salivary proteins in the plant host was provided, which will enable further investigation on the function of salivary protein in plant hosts.
This protocol provides techniques to detect the salivary proteins of leafhoppers with piercing-sucking mouthparts by collecting the feeding plants. Some remarkable points should be noted to obtain the best and reliable results. (1) The viral loading in the infected rice plants is critical. The viral titers in rice plants directly affect the acquisition of insects. When the insect colony is highly viruliferous, the probability of collecting viral proteins in saliva in vitro will be increased. Therefore, the viral proteins released from the saliva to the plant host will be much easier to detect. Choosing infected plants that display significant symptoms is recommended to serve as the source for viruses. (2) Detection timing. It is believed that some of the salivary proteins are transient in the plant host because they are subjected to degradation by the plant host or diluted in the plant. Instant detection of the feeding plants after the 2-day feeding is recommended. It is also hypothesized that a longer retention time would be better for some specific salivary proteins in the plant. Therefore, the detection timing of some salivary proteins could be determined in further studies. (3) Ensure that there are enough replicates. The number of insects that survive will decrease because the confined insects in the feeding cage have limited activity and ability to feed. Using enough replicates will help to enrich the salivary proteins. Three to five replicates are typically enough. If there is only one replicate, it would be better to confine 15-20 leafhoppers to feed on one rice seedling.
These representative results showed the presence of viral protein P8 in the feeding plant of viruliferous leafhoppers. It was revealed that the virus mixed with the flow of saliva is released from the salivary glands. It was then released from the insect to the plant host, ultimately finishing the viral horizontal transmission. However, it is still unknown whether Vg plays the role of elicitor or effector in the process of insect feeding and whether or not it triggers or suppresses the immune response of the plant host. Previously, the saliva of more than 10,000 R. dorsalis was collected via membrane feeding and analyzed using LC-MS/MS (unpublished data). The presence of Vg in the saliva was verified, although its function is unknown. Here, it has been proven that Vg also exists in the saliva of N. cincticeps and is even released to the plant. Combined with the studies on planthoppers16, it is presumed that the presence of Vg in hemipteran saliva is universal. A new finding reports that the Vg of planthopper saliva is an effector to damage the plant defense system34. More studies are required to address whether the Vg in most hemipteran saliva functions as an effector. It is believed that this protocol provides a stable and reliable methodology to examine the presence of salivary proteins secreted by leafhoppers in plants. This protocol is expected to be applicable for the detection of salivary proteins of most hemipterans.
Divulgations
The authors have nothing to disclose.
Acknowledgements
This work was supported by grants from the National Natural Science Foundation of China (31772124 and 31972239) and Fujian Agriculture and Forestry University (Grant KSYLX014).
Materials
| Reagents | |||
| Tris base | Roche | D609K69032 | For 5×Tris-glycine buffer and 10×TBS buffer preparation |
| glycine | Sigma-Aldrich | WXBD0677V | For 5×Tris-glycine buffer preparation |
| SDS | Sigma-Aldrich | SLCB4394 | For 5×Tris-glycine buffer preparation |
| NaCl | Sinopharm Chemical Reagent Co., Ltd | 10019318 | For 10×TBS buffer preparation |
| KCl | Sinopharm Chemical Reagent Co., Ltd | 10016318 | For 10×TBS buffer preparation |
| ß-mercaptoethanol | Xiya Reagent | B14492 | For 4× protein sample buffer preparation |
| bromophenol blue | Sigma-Aldrich | SHBL3668 | For 4× protein sample buffer preparation |
| glycerol | Sinopharm Chemical Reagent Co., Ltd | 10010618 | For 4× protein sample buffer preparation |
| methanol | Sinopharm Chemical Reagent Co., Ltd | 10014118 | For transfer buffer preparation |
| Tween 20 | Coolaber SCIENCE&TeCHNoLoGY | CT30111220 | For TBST preparation |
| non-fat dry milk | Becton.Dickinso and company | 252038 | For membrane blocking, antibodies dilution |
| goat anti-rabbit IgG | Sangon Biotech | D110058-0001 | Recognization of the primary andtibody |
| ECL Western kit | ThermoFisher Scientific | 32209 | Chemiluminescent substrate |
| nitrocellulose membrane | Pall Corporation | 25312915 | For proteins transfer |
| Buffers and Solutions | |||
| Buffer | Composition | Comments/Description | |
| 5×Tris-glycine buffer | 15.1 g Tris base 94 g glycine 5 g SDS in 1 L sterile water |
Stock solution | |
| 1×Tris-glycine buffer | 200 mL of 5×Tris-glycine buffer 800 mL sterile water |
Work solution, for SDS-PAGE | |
| 10×Tris-buffered saline (TBS) buffer | 80 g NaCl 30 g Tris base 2 g KCl in 1 L sterile water |
Stock solution | |
| TBS with Tween 20 (TBST) solution | 100 mL 10×TBS solution 3 mL Tween 20 900 mL sterile water |
Work solution | |
| 4× protein sample buffer | 8 g SDS 4 mL ß-mercaptoethanol 0.02 g bromophenol blue 40 mL glycerol in 40 mL 0.1 M Tris-HCl (pH 6.8) |
For protein extraction | |
| Transfer buffer | 800 mL Tris-glycine buffer 200 mL methanol |
For protein transfer | |
| Instruments | |||
| Bromophenol blue | Sigma-Aldrich | SHBL3668 | For 4x protein sample buffer preparation |
| Constant temperature incubator | Ningbo Saifu Experimental Instrument Co., Ltd. | PRX-1200B | For rearing leafhoppers |
| Electrophoresis | Tanon Science & Technology Co.,Ltd. | Tanon EP300 | For SDS-PAGE |
| Electrophoretic transfer core module | BIO-RAD | 1703935 | For SDS-PAGE |
| glycerol | Sinopharm Chemical Reagent Co., Ltd | 10010618 | For 4x protein sample buffer preparation |
| glycine | Sigma-Aldrich | WXBD0677V | For 5x Tris-glycine buffer preparation |
| goat anti-rabbit IgG | Sangon Biotech | D110058-0001 | Recognization of the primary andtibody |
| High-pass tissue grinding instrument | Shanghai Jingxin Industrial Development Co., Ltd. | JXFSIPRP-24 | For grinding plant tissues |
| KCl | Sinopharm Chemical Reagent Co., Ltd | 10016318 | For 10x TBS buffer preparation |
| methanol | Sinopharm Chemical Reagent Co., Ltd | 10014118 | For transfer buffer preparation |
| Mini wet heat transfer trough | BIO-RAD | 1703930 | For SDS-PAGE |
| NaCl | Sinopharm Chemical Reagent Co., Ltd | 10019318 | For 10x TBS buffer preparation |
| nitrocellulose membrane | Pall Corporation | 25312915 | For proteins transfer |
| non-fat dry milk | Becton.Dickinso and company | 252038 | For membrane blocking, antibodies dilution |
| Pierce ECL Western kit | ThermoFisher Scientific | 32209 | Chemiluminescent substrate |
| Protein color instrument | GE Healthcare bio-sciences AB | Amersham lmager 600 | For detecting proteins |
| SDS | Sigma-Aldrich | SLCB4394 | For 5x Tris-glycine buffer preparation |
| Tris base | Roche | D609K69032 | For 5x Tris-glycine buffer and 10×TBS buffer preparation |
| Tween 20 | Coolaber SCIENCE&TeCHNoLoGY | CT30111220 | For TBST preparation |
| Vertical plate electrophoresis tank | BIO-RAD | 1658001 | For SDS-PAGE |
| Water bath | Shanghai Jinghong Experimental equipment Co., Ltd. | XMTD-8222 | For boil the protein samples |
| β-mercaptoethanol | Xiya Reagent | B14492 | For 4x protein sample buffer preparation |
References
- Hogenhout, S. A., Ammar el, D., Whitfield, A. E., Redinbaugh, M. G. Insect vector interactions with persistently transmitted viruses. Annual Review of Phytopathology. 46, 327-359 (2008).
- Cranston, P. S., Gullan, P. J., Resh, V. H., Carde, R. T. Phylogeny of insects. Encyclopedia of Insects. , (2003).
- Ammar el, D., Tsai, C. W., Whitfield, A. E., Redinbaugh, M. G., Hogenhout, S. A. Cellular and molecular aspects of rhabdovirus interactions with insect and plant hosts. Annual Review of Entomology. 54, 447-468 (2009).
- Wei, T., Li, Y. Rice reoviruses in insect vectors. Annual Review of Phytopathology. 54, 99-120 (2016).
- Hattori, M., Konishi, H., Tamura, Y., Konno, K., Sogawa, K. Laccase-type phenoloxidase in salivary glands and watery saliva of the green rice leafhopper, Nephotettix cincticeps. Journal of Insect Physiology. 51 (12), 1359-1365 (2005).
- Ma, R., Reese, J. C., William, I. V., Bramel-Cox, P. Detection of pectinesterase and polygalacturonase from salivary secretions of living greenbugs, schizaphis graminum (Homoptera: aphididae). Journal of Insect Physiology. 36 (7), 507-512 (1990).
- Miles, P. W. Dynamic aspects of the chemical relation between the rose aphid and rose buds. Entomologia Experimentalis et Applicata. 37 (2), 129-135 (2011).
- Urbanska, A., Tjallingii, W. F., Dixon, A., Leszczynski, B. Phenol oxidising enzymes in the grain aphid’s saliva. Entomologia Experimentalis et Applicata. 86 (2), 197-203 (1998).
- Miles, P. W., Peng, Z. Studies on the salivary physiology of plant bugs: detoxification of phytochemicals by the salivary peroxidase of aphids. Journal of Insect Physiology. 35 (11), 865-872 (1989).
- Will, T., van Bel, A. Physical and chemical interactions between aphids and plants. Journal of Experimental Botany. 57 (4), 729-737 (2006).
- Ma, R. Z., Reese, J. C., Black, W. C., Bramel-Cox, I. Chlorophyll loss in a greenbug-susceptible sorghum due to pectinases and pectin fragments. Journal of the Kansas Entomological Society. 71 (1), 51-60 (1998).
- Madhusudhan, V. V., Miles, P. W. Mobility of salivary components as a possible reason for differences in the responses of alfalfa to the spotted alfalfa aphid and pea aphid. Entomologia Experimentalis et Applicata. 86 (1), 25-39 (1998).
- Funk, C. J. Alkaline phosphatase activity in whitefly salivary glands and saliva. Archives of Insect Biochemistry & Physiology. 46 (4), 165-174 (2010).
- Hogenhout, S. A., Bos, J. I. Effector proteins that modulate plant-insect interactions. Current Opinion in Plant Biology. 14 (4), 422-428 (2011).
- Tomkins, M., Kliot, A., Maree, A. F., Hogenhout, S. A. A multi-layered mechanistic modelling approach to understand how effector genes extend beyond phytoplasma to modulate plant hosts, insect vectors and the environment. Current Opinion in Plant Biology. 44, 39-48 (2018).
- Huang, H. J., Lu, J. B., Li, Q., Bao, Y. Y., Zhang, C. X. Combined transcriptomic/proteomic analysis of salivary gland and secreted saliva in three planthopper species. Journal of Proteomics. , (2018).
- Hogenhout, S. A., Bos, J. I. Effector proteins that modulate plant–insect interactions. Current Opinion in Plant Biology. 14 (4), 422-428 (2011).
- Sun, P., et al. A mosquito salivary protein promotes flavivirus transmission by activation of autophagy. Nature Communications. 11 (1), 260 (2020).
- Sri-In, C., et al. A salivary protein of Aedes aegypti promotes dengue-2 virus replication and transmission. Insect Biochemistry and Molecular Biology. 111, 103181 (2019).
- Conway, M. J., et al. Aedes aegypti D7 saliva protein inhibits dengue virus infection. Plos Neglected Tropical Diseases. 10 (9), 0004941 (2016).
- Wang, N., et al. A whitefly effector Bsp9 targets host immunity regulator WRKY33 to promote performance. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 374 (1767), 20180313 (2019).
- Omura, T., Yan, J. Role of outer capsid proteins in transmission of phytoreovirus by insect vectors. Advances in Virus Research. 54, 15-43 (1999).
- Chen, Q., Liu, Y., Long, Z., Yang, H., Wei, T. Viral release threshold in the salivary gland of leafhopper vector mediates the intermittent transmission of rice dwarf virus. Frontiers in Microbiology. 12, 639445 (2021).
- Fukushi, T. Further studies on the dwarf disease of rice plant. Journal of the Faculty of Agriculture, Hokkaido Imperial University. 45 (3), 83-154 (1940).
- Miyazaki, N., et al. The functional organization of the internal components of rice dwarf virus. Journal of Biochemistry. 147, 843-850 (2010).
- Wei, T., Shimizu, T., Hagiwara, K., Kikuchi, A., Omura, T. Pns12 protein of rice dwarf virus is essential for formation of viroplasms and nucleation of viral-assembly complexes. Journal of General Virology. 87, 429-438 (2006).
- Chen, Q., Zhang, L., Chen, H., Xie, L., Wei, T. Nonstructural protein Pns4 of rice dwarf virus is essential for viral infection in its insect vector. Virology Journal. 12, 211 (2015).
- Chen, Q., et al. Nonstructural protein Pns12 of rice dwarf virus is a principal regulator for viral replication and infection in its insect vector. Virus Research. 210, 54-61 (2015).
- Chen, Q., Zhang, L., Zhang, Y., Mao, Q., Wei, T. Tubules of plant reoviruses exploit tropomodulin to regulate actin-based tubule motility in insect vector. Scientific Reports. 7, 38563 (2017).
- Wei, T., et al. The spread of Rice dwarf virus among cells of its insect vector exploits virus-induced tubular structures. Journal of Virology. 80 (17), 8593-8602 (2006).
- Mao, Q., et al. Insect bacterial symbiont-mediated vitellogenin uptake into oocytes to support egg development. mBio. 11 (6), 01142 (2020).
- Tufail, M., Takeda, M. Molecular characteristics of insect vitellogenins. Journal of Insect Physiology. 54 (12), 1447-1458 (2008).
- Sappington, T. W., Raikhel, A. S. Molecular characteristics of insect vitellogenins and vitellogenin receptors. Insect Biochemistry and Molecular Biology. 28 (5-6), 277-300 (1998).
- Ji, R., et al. Vitellogenin from planthopper oral secretion acts as a novel effector to impair plant defenses. New Phytologist. , (2021).
