In a recently published work5, we applied the protocol described above to explore the early stages of PDAC development using a mouse model. The mouse was genetically engineered to include the cassettes Ptf1a–CreER, LSL-Kras-G12D, LSL-tdTomato7, which allow the expression of constitutively active KRAS in acinar cells after tamoxifen injection.
After cervical dislocation (according to the mouse ethical protocol), the pancreas was resected, and the protocol described above was applied (see Figure 1 and Protocol).
To optimize the dissociation protocol, the dissociation was performed with or without pre-incubation of the tissue with trypsin C. It was found that pre-incubation with trypsin C accelerated the dissociation without affecting the viability. In addition, different collagenases, including collagenase D, collagenase 1a, and collagenase P, were tried (see Table 1). It was found that using collagenase D resulted in massive cell death, and indeed in other studies that used this collagenase, the fraction of acinar cells was very small8. The use of collagenase 1a also did not provide the expected results, as even after a 90 min incubation with collagenase 1a, the tissue was not dissociated. Only collagenase P allowed us to dissociate the tissue and maintain high viability of all the cell types.
These experiments were repeated at seven different time points post-tamoxifen injection. Simultaneously with acinar-to-ductal metaplasia and PanIN lesion formation, there was an accumulation of stromal and immune cells, including fibroblasts, and the tissue became desmoplastic and stiff. The different time points post-tamoxifen injection allowed us to examine the protocol under several different tissue states and measure the recovery of epithelial, stromal, and immune cells.
A photo of a mouse is shown in Figure 2A and the isolated pancreas, a fluffy tissue, is shown in Figure 2C. The red color of the tissue results from the expression of tdTomato in the acinar cells. The protocol was used successfully with all the tissue states and with human samples. However, incubation times with trypsin and collagenase may vary. It is therefore important to monitor the sample and observe the dissociation of the tissue and cell viability every few minutes under the microscope. At an early stage of the dissociation protocol, there were low numbers of isolated cells and large numbers of clumps (Figure 2D, left). Our aim was to achieve isolated viable cells, as shown in Figure 2D, middle, and avoid a reduction in the number of viable cells (Figure 2D, right).
It is important to note that longer incubation times reduced the viability of the cells. Starting from a tissue 0.5 x 10 cm3 in size, we recovered a total of 5 x 106 cells, 5 x 105 of which were tdTomato positive cells. According to the fluorescence-activated cell sorting (FACS) analysis in Figure 3, our pancreas dissociation protocol supports the recovery of multiple cell types, including acinar cells, ductal cells, endothelial cells, fibroblasts, immune cells, and pericytes with high viability. However, the actual ratio between the number of cells from each type in the tissue before dissociation may be different, inferred from the fluorescence images of pancreatic frozen sections that include tdTomato positive cells (Figure 2E). The high viability achieved using the protocol detailed above is shown by the FACS analysis (Figure 3) and the live cell/dead cell counting done using a hemocytometer (Figure 3H).
For each sample, we performed scRNA-seq and, consistent with the FACS analysis, we confirmed the detection of all the above-mentioned cell types (Figure 4A,B), in each of the resected tissue from all the time points post-tamoxifen injection. We also examined the expression of carboxypeptidase 1 (Cpa1), which encodes carboxypeptidase1. Cpa1 expression in acinar cells is extremely high and can indicate to what extent acinar cells were lysed during the dissociation, as well as the extent to which they have contaminated the transcriptome of other cell types. We detected minimal contamination, as can be seen in Figure 4C,D. The gentle dissociation also results in high viability that allows us to follow-up with cell sorting of desired cell types for additional experiments (Figure 3).
Together, the quality of the dissociation protocol supports different follow-up experiments, including scRNA-seq.
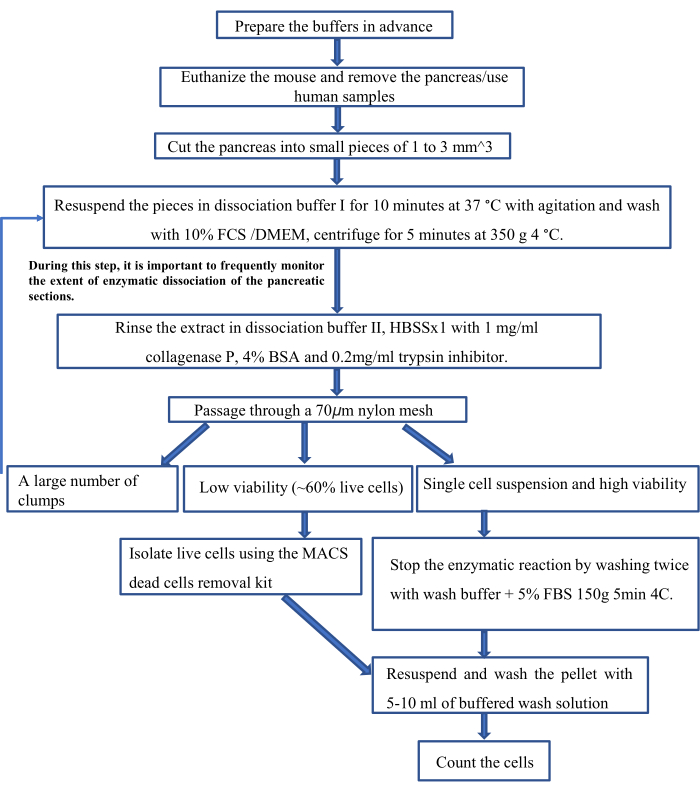
Figure 1: Scheme of the protocol. Please click here to view a larger version of this figure.
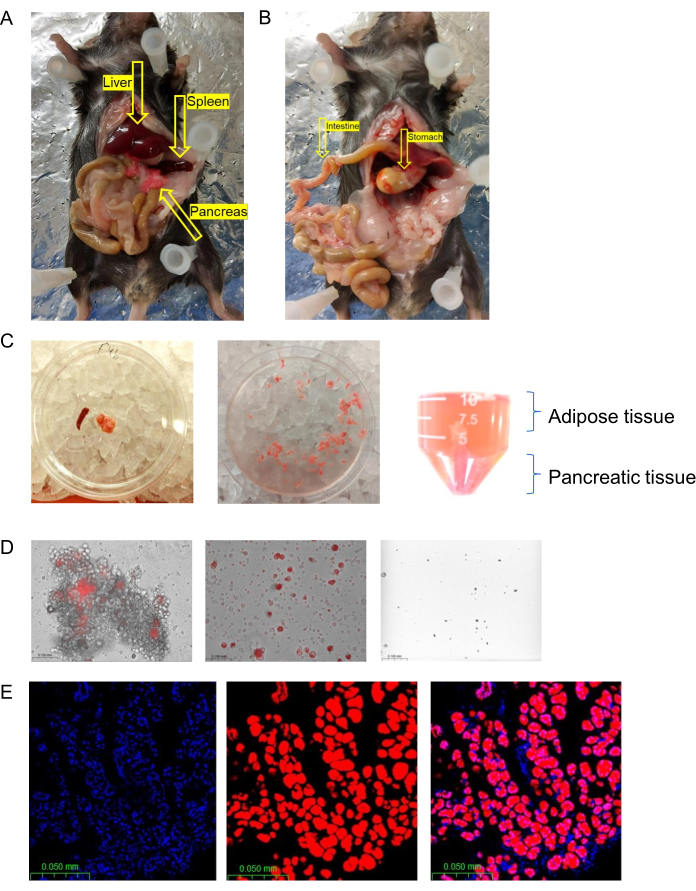
Figure 2: Pancreatic cell isolation. (A) A photo showing the pancreas (in red), liver, and spleen of the mouse after fully opening the abdominal cavity. (B) The same mouse as in (A) after removal of the pancreas. (C) The dissection of the pancreas. The pancreas after removal from the animal (left). The pancreas after cleavages into small pieces (middle). Phase separation after centrifugation (right). (D) Monitoring the cells under the microscope to examine single-cell viability. Total primary pancreatic cell isolation (20x). Clumps of cells after 15 min of enzymatic reaction (left). Single-cell suspension after 25 min of enzymatic reaction (middle). Reduced cell viability after a long incubation (right). (E) Fluorescence images of a frozen pancreatic section. Acinar cells are tdTomato positive; DAPI staining in blue (photo at 40x). Please click here to view a larger version of this figure.
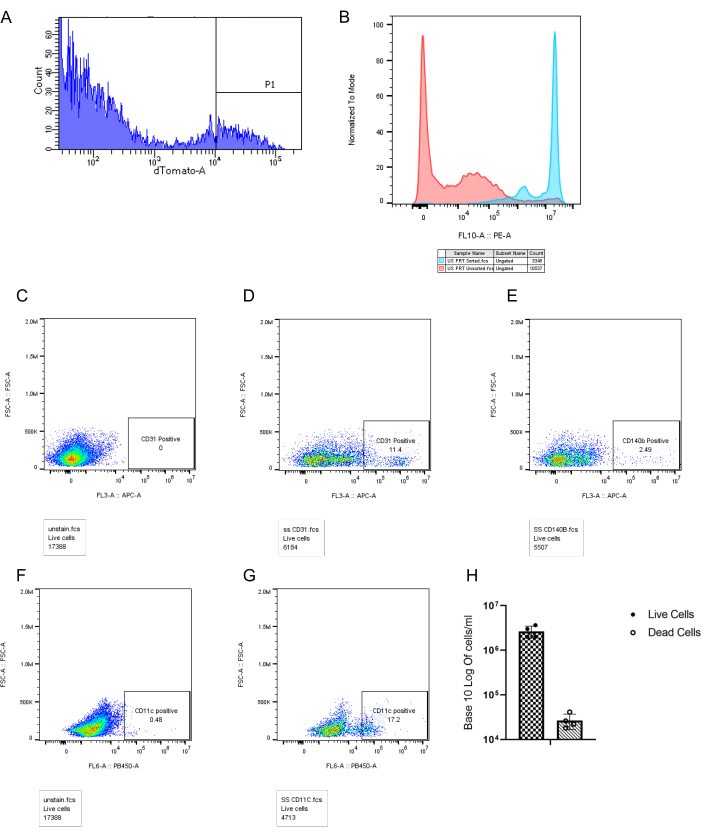
Figure 3: FACS analysis of single-cell suspension of pancreatic cancer. (A) tdTomato positive and negative cells. (B) Flow cytometry of unsorted (red) versus sorted (blue) acinar cells. (C,F) Unstained cells, APC and PB450. (D) FACS analysis after staining with Anti-CD31, a marker of endothelial cells. (E) FACS analysis after staining with Anti-CD140b, a marker of pericytes. (G) FACS analysis after staining with Anti-CD11c, a marker of dendritic cells and macrophages. (H) Ratio of live cells and dead cells in four different pancreas cell isolations. Please click here to view a larger version of this figure.
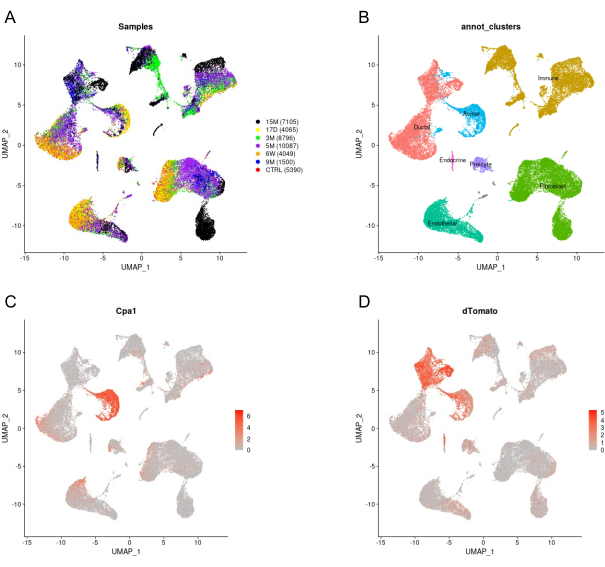
Figure 4: Analysis of scRNA-seq data that was produced using the protocol that we describe in the current article. (A) UMAP showing scRNA-seq of a mouse pancreas at different time points post-tamoxifen injection, as indicated in the right side of the panel. (B) Cell types were determined based on known markers. (C,D) Cells are colored according to the expression level of Cpa1 (in C) or the level of tdTomato (in D). Please click here to view a larger version of this figure.
