The results in Fig. 2 show percentages and absolute numbers of different BSL populations recovered from brains of perfused or unperfused malaria-infected and naïve control mice. Isolated BSL were stained with PE-anti-NK1.1 and APC-anti-TCR-β antibodies as indicated in the Protocol text. Consistent with previous findings 7-9, αβTCR+ T cells comprised a high proportion of the BSL pool in brains of perfused malaria-infected mice (day 6 p.i). This population appeared to be significantly underrepresented in brains of non-perfused animals (Fig. 2A-B). The apparent reduction of T cell frequencies in non-perfused brains was associated with a considerably higher percentage and total number of double negative cells (αβTCR– NK1.1–) in these animals (Fig. 2A-B). High percentage and numbers on double negative cells in unperfused brains were not only detected in malaria-infected mice but also in naïve controls, suggesting that these cells are non-inflammatory leukocytes present in brain blood vessels that are recovered together with inflammatory cells if intracardial perfusion is not performed.
Two further examples of BSL analysis are shown in Fig. 3 and Fig. 4. Briefly, C57BL/6 mice were infected with P. berghei ANKA (1×106 pRBC). Temporal associations between leukocyte recruitment to the brain and the onset of neurological symptoms (between days 6-8 p.i) have been demonstrated in P. berghei ANKA infected mice 7-9. In this particular example, malaria-infected mice were euthanized on day 6 p.i, and the BSL were purified as described in the Protocol text. Isolated BSL were stained with PE-anti-NK1.1, APC-anti-TCR-β, FITC-anti-CD4 and PerCPCy5.5-anti-CD8α antibodies. The samples were acquired using a BD Biosciences FACSCalibur flow cytometer and the analysis performed using Weasel 3.0.1 software. Viable cells were gated by forward and side scatter. The percentages of NK cells, T lymphocytes and NKT cells (NK1.1+TCR+ cells) sequestered in brain of naïve and malaria-infected mice (day 6 p.i) are indicated in the top panels (Fig. 3A). The bottom panels show percentages of CD4+ and CD8+ T lymphocytes amongst gated NK1.1–αβTCR+ cells (Fig. 3A). Fig. 3B depicts an example in which absolute numbers of CD4+ and CD8+ brain-sequestered T cells were calculated on day 6 p.i with P. berghei ANKA. In general, 20,000-100,000 BSL can be recovered from brains of P. berghei ANKA infected mice. Several factors such as time p.i, susceptibility of the mouse strain or inclusion of anti-inflammatory treatments might influence cell recovery. Routinely, one individual brain provides enough cells for 1 set of antibody stainings.
In the example provided in Fig. 4 the chemokine receptor usage of brain-sequestered T lymphocytes was analyzed. Mice were infected with P. berghei ANKA and BSL were isolated on day 6 p.i. Cells were stained with APC-anti-TCR-β, PE-anti-CXCR3 and biotinylated-anti-CCR5 antibodies, followed by incubation with a streptavidin-PerCPCy5.5 conjugate. Samples were acquired and analyzed as per Fig. 3. The histograms represent percentages of αβTCR+ cells amongst total BSL in malaria-infected and naïve control mice. The contour plots in the middle and bottom panels show the percentages of CXCR3+ and CCR5+ cells within gated αβTCR+ cells.
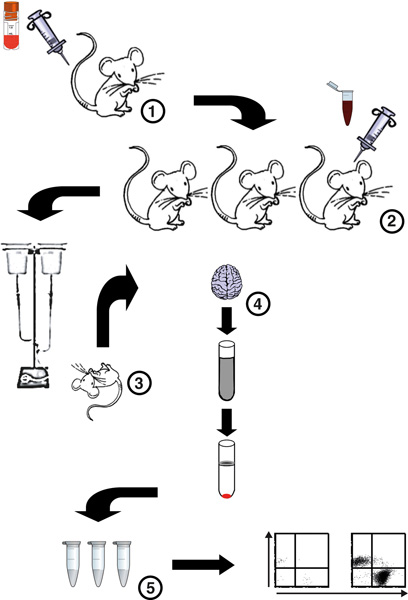
Figure 1. Flow chart of the procedure for isolation and analysis of BSL from P. berghei ANKA-infected mice. (1) A cryopreseved aliquot of P. berghei ANKA pRBC is defrosted and 1 or 2 donor mice are infected. (2) Four days later parsitemia levels of donor mice are calculated, donor mice are bled and blood dilutions prepared for infection of experimental mice. A malaria infection is then set up in severe malaria-susceptible C57BL/6 mice. (3) On days 5-7 p.i, mice are euthanized by CO2 inhalation and immediately intracardially perfused to remove all circulating non-adherent cells. (4) Brains are then harvested and a brain homogenate prepared by digestion of the tissue with Collagenase D and DNAse I. BSL are then separated from myelin and cell debris by a Percoll gradient. (5) Precipitated BSL are then washed, counted using a hemocytometer, stained with fluorescent antibodies and analyzed by flow cytometry.
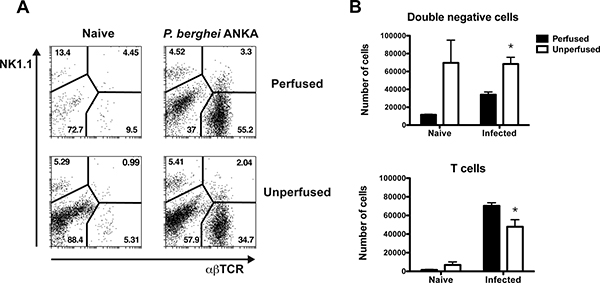
Figure 2. BSL analysis in brains of perfused or unperfused mice. C57BL/6 mice were infected with P. berghei ANKA (1×106 pRBC). Brains were harvested on day 6 p.i from perfused or unperfused animals. The BSL were isolated, stained with anti-NK1.1 and anti-TCR fluorescent antibodies and analyzed by flow cytometry. (A) Dot plots depict percentages of NK cells, T lymphocytes, NK1.1+TCR+ cells and double negative cells NK1.1–TCR– in perfused (top panels) or unperfused (bottom panels) malaria-infected and naïve control mice. (B) The absolute number of double negative cells and T lymphocytes and was calculated on day 6 p.i with P. berghei ANKA and naïve control mice. Each bar represents the mean of 3 samples ± SEM, *p>0.05.
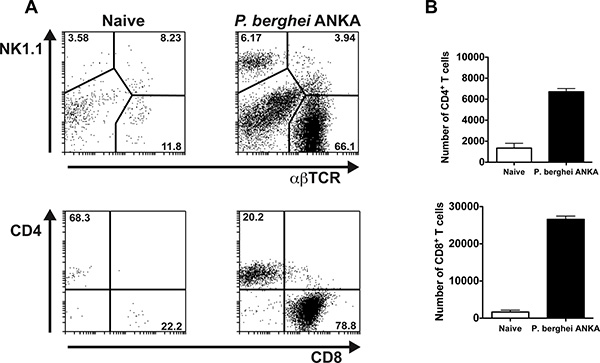
Figure 3. NK cells and T lymphocytes can be detected in brains of P. berghei ANKA infected mice. C57BL/6 mice were infected with P. berghei ANKA (1×106 pRBC). Brains were extracted on day 6 p.i after perfusion of the euthanized animals. The BSL were isolated, stained with anti-NK1.1, anti-TCR, anti-CD4 and anti-CD8 fluorescent antibodies and analyzed by flow cytometry. (A) Top panels depict percentages of NK cells, T lymphocytes and NK1.1+TCR+ cells in malaria-infected and naïve control mice. Bottom panels illustrate percentages of CD4+ and CD8+ T cells within gated αβTCR+ NK1.1– cells. Representative dot plots are shown. (B) The absolute number of brain-sequestered CD4+ and CD8+ T cells was calculated on day 6 p.i with P. berghei ANKA and naïve control mice. Each bar represents the mean of 3 samples ± SEM.
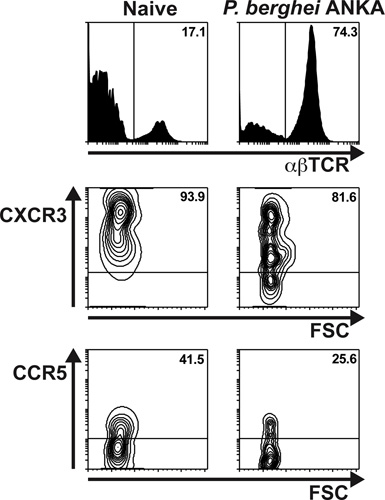
Figure 4. The majority of T lymphocytes migrating to the brain in response to severe malaria infection express the chemokine receptor CXCR3. C57BL/6 mice were infected with P. berghei ANKA (1×106 pRBC). Brains were extracted on day 6 p.i after extensive perfusion of the euthanized animals. The BSL were isolated, stained with anti-TCR, anti-CXCR3 and anti-CCR5 antibodies and analyzed by flow cytometry. Top panels depict percentages of αβT lymphocytes in malaria-infected and naïve control mice. The contour plots illustrate percentages of CXCR3+ (middle panels) and CCR5+ (bottom panel) cells within gated αβTCR+ lymphocytes.
| Problem | Troubleshooting |
| Incomplete perfusion |
|
| Not enough BSL recovered |
|
Table 1.
