In this study, we performed whole-mount immunostaining on embryos of asexual pea aphids (Figure 1A). These females produce offspring parthenogenetically and viviparously. These female embryos develop within egg chambers of the ovarian tubules (ovarioles) (Figure 1B and Figure 2A). Before microscopy, the dissected ovarioles are the staining targets; however, separation of egg chambers is required for observation of embryos under a microscope (Figure 2B–D).
Increase of tissue permeability
Proteinase K (PK) treatment is a standard approach for enhancing tissue permeability, but for embryos of some model organisms-such as Caenorhabditis elegans (nematode), Drosophila melanogaster (fly), and Danio rerio (zebrafish)-this step is optional. In the pea aphid, the requirement for PK treatment is stage-dependent: for germaria and embryos prior to gastrulation (stages 0-7), PK treatment can be omitted; but for embryos under germband extension (stage 11) or in later stages this step is highly recommended. For example, during mid embryogenesis signals were barely detected in embryos without PK treatment (Figure 3A–C). By contrast, signal intensity was significantly enhanced in embryos subjected to PK digestion (Figure 3A'–C', A"–C").
Reduction of background staining
A high level of the endogenous peroxidase (POD) activity was identified in the embryonic tissues of aphids. To suppress this enzyme activity, the paraformaldehyde-fixed embryos were incubated in the presence of hydrogen peroxide (H2O2), a common reagent for the oxidation of POD. Our results showed that H2O2 treatment did not suppress the activity of endogenous POD effectively in the late-stage embryos (Figure 4A, D). By contrast, background staining was largely reduced in embryos subjected to methanol incubation (Figure 4B, C, E, F). Before application of the primary antibody embryos were blocked in solution containing NGS and BSA as described in standard protocols for antibody staining. However, residual background staining in the aphid embryos was detected (Figure 3A'–C'). This problem was resolved after replacing NGS/BSA with the blocking reagent supplied by a buffer set for DIG-labeling experiments such as in situ hybridization (Figure 3A"–C"). We thus conclude that post-fixation with methanol and incubation with the DIG-B blocking reagent are both essential for reducing background staining in the aphid embryos.
PK treatment and the DIG-B blocking reagent are required not only for effective signal detection using the chromogenic but also fluorescent approaches. Actin staining with phalloidin or staining for methanol-sensitive epitope, however, does not work in embryos subjected to pre-fixation with methanol, which can destroy the native form of actin or antibodies. Accordingly, this treatment should be avoided when performing fluorescence immunostaining. Apart from eliminating the methanol treatment steps immunofluorescence allows multi-labeling experiments in the aphid embryos. Using confocal microscopy, for example, pea aphid Vasa1 protein (ApVas1) in germ cells, α-tubulin in microtubules, F-actin in microfilaments, and DNA in nuclei could be concurrently marked and visualized (Figure 5A, C). In comparison with the chromogenic method (Figure 5B), confocal sectioning of fluorescently labeled samples provides better resolution of localized signals such as ApVas1 in the germ plasm and cellularizing germ cells in early aphid embryos (Figure 5C', D–D"). Adopting the conditions for labeling germ cells described above, the Engrailed/Invected protein in segments of the extending germ band was also stained with the antibody 4D9 (Figure 6). This shows that our protocol for immunostaining-including chromogenic and fluorescent methods-can be effectively applied to signal detection in both germline and somatic cell lineages in aphids.
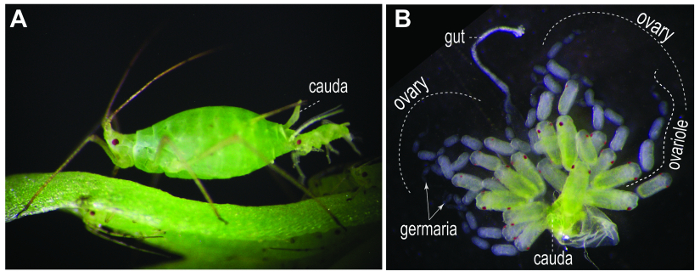
Figure 1. Parthenogenetic viviparous pea aphids. (A) A first-instar nymph emerging from an asexual viviparous female adult. (B) A pair of dissected ovaries. Each ovary is composed of 7 ovarian tubules (ovarioles). Nonetheless, the number of ovarioles within an ovary may be variable between strains12. An ovariole contains a germarium in the tip (arrows), 1-2 oocytes, and 5-7 embryonic chambers. Gut and cauda are usually associated with the dissected ovaries. Please click here to view a larger version of this figure.
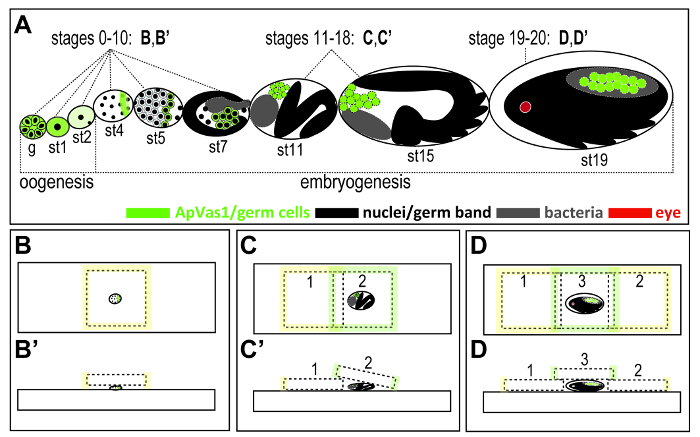
Figure 2. Illustration of strategies for mounting aphid embryos at different stages of development. (A) Outline of embryonic development in the ovariole dissected from a parthenogenetic viviparous female adult. Brief oogenesis (germarial stage and stages 0-2) is followed by embryogenesis (stages 3-20). Outline of embryogenesis: formation of syncytial blastoderm (stages 3-5); blastulation (stages 6-7); gastrulation (stages 8-10); elongation of germ band (stages 11-14); katatrepsis (stages 15); post katatrepsis and germ band retraction (stages 16-17); organogenesis (stages 18-20). Color keys are at the bottom of the figure. (B, B') Germarium and stages 0-10 embryos. No coverslip bridge is required. (C, C') Stages 11-18 embryos. A coverslip bridge is required. (D, D') Stages 19-20 embryos. Double coverslip bridges are required. Rolling the embryos by sliding the coverslip can create different angles of observation. Sizes of coverslips: 22 x 22 mm in (B), 18 x 18 mm in (C) and (D). The thickness of coverslips: 0.13 to 0.16 mm. Embryonic staging followed Miura et al.12 Abbreviations: g: germarium; st: stage. Please click here to view a larger version of this figure.
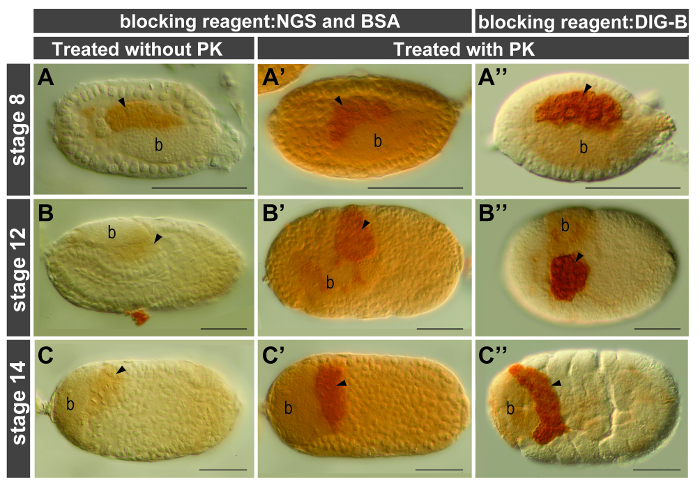
Figure 3. Proteinase K treatment and comparison of reagents with different blocking effects. Anterior of egg chambers is to the left; all views are lateral except embryos shown in (B", C', C"), which are dorsal. Embryos are all stained using ApVas1 antibody (dilution 1:500) and signals of ApVas1 are developed within 10-20 sec. Arrowheads indicate location of germ cells. (A–C) Embryos without treatment of proteinase K (PK). ApVas1 signals in germ cells are barely detected. (A'-C', A"-C") Comparison of background staining in embryos blocked with NGS and BSA (A'-C') and of the commercial blocking reagent in the DIG Wash and Block Buffer Set (DIG-B) (A"-C"). Background is significantly reduced in embryos shown in (A"-C"). Abbreviation: b: bacteria. Scale bars: 100 µm. Please click here to view a larger version of this figure.
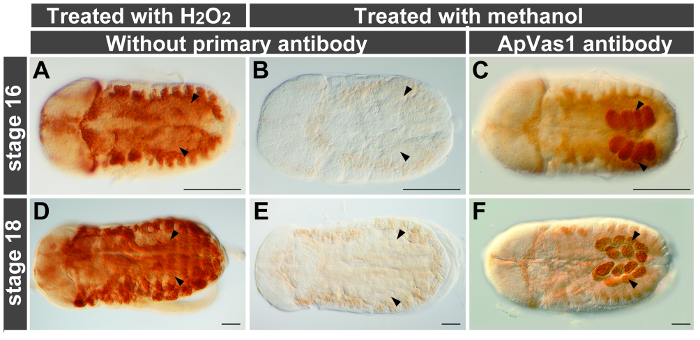
Figure 4. Minimizing background staining with methanol. Anterior of embryos is to the left; all views are dorsal. Embryos are treated with PK for increasing permeability of antibody. Arrowheads indicate location of germ cells. (A, B, D, E) Comparison of treatments with hydrogen peroxide (H2O2) and methanol. Embryos are only stained with secondary antibody. H2O2 treatment (0.3% w/v, 10 min): high background (A, D); methanol treatment (100%, 60 min): low background (B, E). (C, F) Primary antibody staining on embryos treated with methanol. Conditions of methanol treatment are identical to those used for embryos shown in (B, E). The ApVas1 antibody preferentially labels the embryonic germ cells. Scale bars: 100 µm. Please click here to view a larger version of this figure.
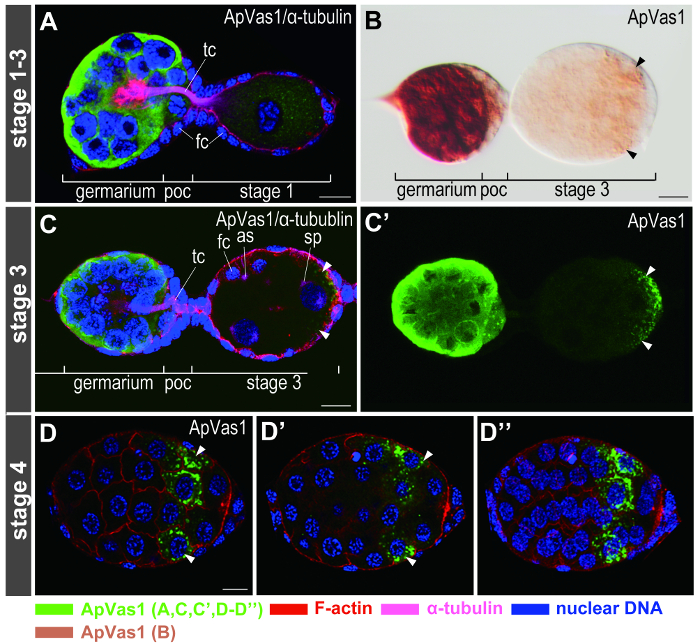
Figure 5. Immunofluorescence staining on early embryos. Anterior of egg chambers is to the left. Dilution ratios of ApVas1 antibody are 1:50 in (A, C–D) and 1:500 in (B). (A) Staining signals, which include ApVas1, α-tubulin, F-actin, and nuclear DNA, are detected using four channels with different wavelengths. Color keys of signals are shown at the bottom of the figure. (B) DIC image of a chromogenic result for comparison. ApVas1 is enriched within the germarium whereas the contrast intensity of posterior localization of ApVas1 (arrowheads) in the stage-3 embryo is not as clear as that shown in (C, C'). (C, C') Enrichment of ApVas1 signals in the egg posterior. Signals localized to the posterior region of the egg chamber (arrowheads) are enhanced by image stacking. Because signals of F-actin and α-tubulin partially mask those of ApVas1 in the posterior (C), an image produced by single-channeled scanning is shown in (C'). (D–D'') Confocal sectioning of ApVas1 localized in the posterior region of the stage-4 embryo. Images shown in (D) and (D') are ApVas1 detected in surface and central focal planes, respectively. (D") is the merged image of all sections from the same embryo as (D) and (D'). Abbreviations: as: aster; fc, follicle cells; poc, prospective oocyte; sp: spindle; tc, trophic cords. Scale bars: 10 µm. Please click here to view a larger version of this figure.
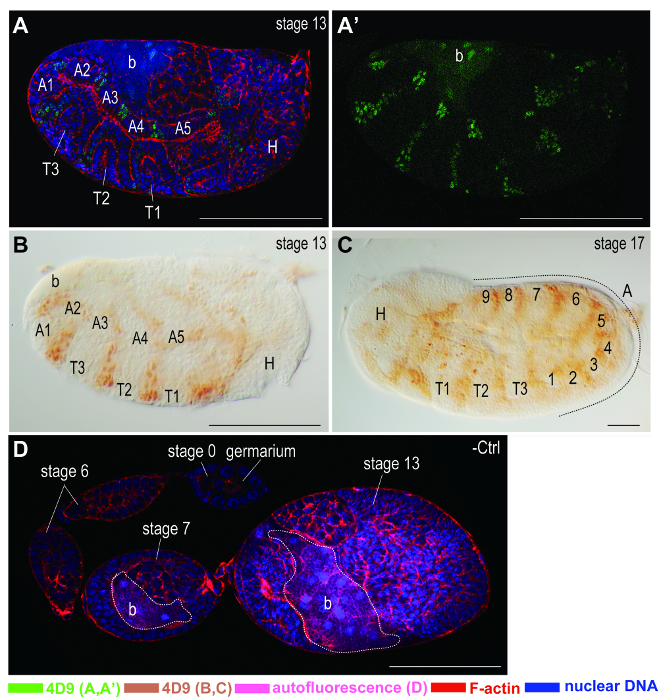
Figure 6. Immunostaining on embryonic segments. Anterior of egg chambers is to the left. PK-treated embryos are stained with the monoclonal antibody 4D9 against Engrailed/Invected, which are expressed in embryonic segments. Color keys of signals are shown at the bottom of the figure. (A, A') Immunofluorescence staining on a stage-13 embryo. (A), a confocal image stacked from images of Engrailed/Invected, α-tubulin, F-actin, and nuclear DNA stainings. (A') a confocal image showing the staining of Engrailed/Invected only. Without the interference of signals from other channels, signals are better displayed. (B, C) Chromogenic staining on embryos at stages 13 and 17 of development, respectively. (B) Stage-13 embryo. (C) Stage-17 embryo. From stages 13 to 17, the growing numbers of segments in the abdomen are labeled by 4D9. (D) Negative control (-Ctrl) staining only with secondary antibody conjugated with Alexa Fluor 633. Almost no immunostaining signals are detected on ovariole without primary antibody. However, autofluorescence of bacteria (dash line) within stage 7 and 13 of egg chambers are detected at the excitation wavelength 488 nm. Abbreviations: A: abdomen; b: bacteria; H: head; T: thorax. Scale bars: 100 µm. Please click here to view a larger version of this figure.
