心肌细胞DNA的合成和倍性的同时评价:一个方法来帮助心肌细胞再生和营业额的量化
Summary
Quantification of cardiomyocyte turnover is challenging. The protocol described here makes an important contribution to this challenge by enabling accurate and sensitive quantification of neo-cardiomyocyte nuclei generation and nuclei ploidy.
Abstract
虽然公认的事实是心脏有潜力有限再生正常老化过程中损伤后心肌细胞和心肌细胞的营业额水平低发生,这些事件的定量仍然具有挑战性。这部分是由于该方法的稀有和事实,即多个蜂窝源向心肌维修。此外,心肌细胞内的DNA复制往往导致多倍体心肌和很少导致通过细胞分裂新的心肌细胞。为了准确地量化这些过程之间的心肌细胞周转歧视是必不可少的。这里描述的协议采用长期核苷的标签,以标示它已出现的利用核隔离和随后的PCM1免疫标记识别DNA复制和心肌细胞核的结果全部核。一起这允许C的核苷标签的准确和灵敏的识别ardiomyocyte核人口。此外,4',6-二脒基-2-苯基标签和细胞核倍性的分析,使新的心肌细胞核的从中多倍体中已掺入的核苷细胞核的鉴别。虽然这种方法不能用于心肌binucleation控制,它允许新的心肌细胞核的快速和强大的量化而占多倍体。这种方法具有许多下游应用包括评估潜在的治疗剂,以提高心肌再生或调查心脏疾病的心肌周转和倍性的影响。该技术也与附加的下游免疫组织学技术,允许核苷掺入定量在所有的心脏细胞类型相容。
Introduction
近年来出现了质疑的证据的假设,心脏是终末分化,有丝分裂后的器官1,2的积累。然而,心肌营业额和再生的量化仍然具有挑战性。
在使用标准的免疫组化技术精确识别罕见的心肌产生的困难以及报告3。此外,心肌细胞产生的细胞来源仍然不明朗与由心肌细胞增殖以及由干细胞分化4-6作出贡献的证据。因此,使用其中所需要的心肌祖细胞表型的知识谱系追踪模式是不可能的,并在一个单一的人口,包括心肌细胞增殖的定量,是不合适的。此外心肌细胞具有无有丝分裂为endoreplication的电位(导致多倍体车diomyocyte)或在没有胞质(导致双核心肌)7,8的有丝分裂。心肌营业额的准确定量依赖于这些事件和真正的新的心肌细胞生成之间分辨能力。这产生独特的并发症,因为DNA复制和细胞周期蛋白依赖性激酶的表达在心肌细胞不完全证明真正细胞分裂9,10。
为了协助新心肌生成的量化,如由贝格曼等 7,11与长期的新颖方法中所述,我们已经联合建立核隔离技术,和pericentriolar材料1(PCM1),用于心肌细胞核识别的免疫标记DNA标记和倍体分析。 PCM-1是积聚在分化的,非循环细胞的核表面上的中心体蛋白质。以前的研究已经表明,对抗体PCM-1特异性标记心肌细胞核7,11,因此PCM1已被用于由多个独立组来识别心肌1,12,13。此外,我们已经表明,PCM1表达映射到肌钙蛋白-CRE转基因小鼠模型14( 补充图1)遗传标记的心肌细胞核。
这里所描述的协议使得不论该细胞起源( 图1A和B),而同时排除核苷标签由于从分析( 图1C和D)多倍体新心肌细胞核产生的在小鼠心脏的精确和灵敏的识别。虽然这种方法不能用于心肌binucleation控制,它允许这是需要的心肌细胞周转的精确定量新心肌细胞核的快速和强大的定量。此外,它提供了一个快速筛选工具,以评估在心肌细胞产生的动态电位变化。
而DNA标记通常涉及将5-溴-2'-脱氧尿苷(BrdU)标记作为胸苷类似物,这里所描述的协议使用,因为它需要一个更迅速更少的处理步骤的5-乙炔基-2'-脱氧尿苷(EDU)基于测定通过量和不需要的DNA进行免疫检测的变性,使它与其他免疫染色的协议,从而增加了该方法的潜在的下游应用相兼容。
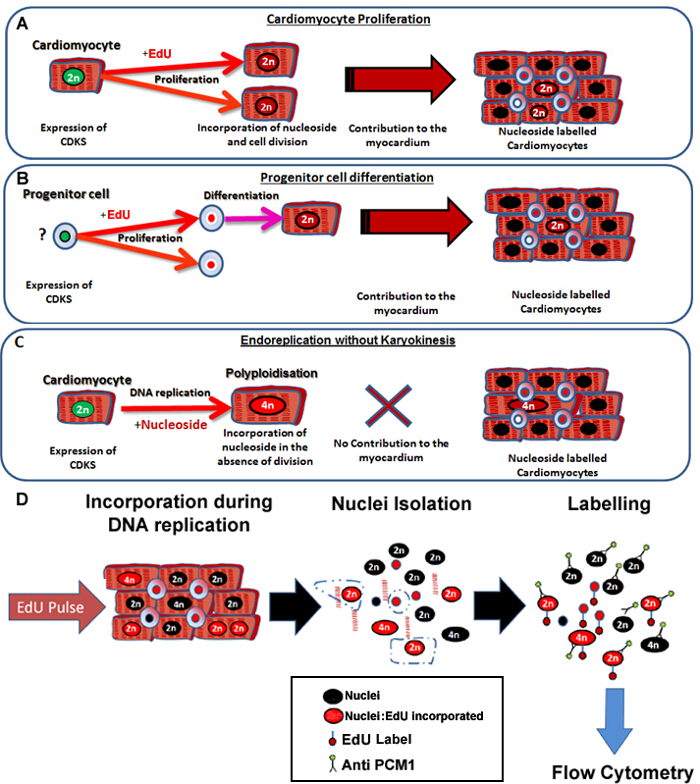
图1: 连续脉冲用EdU标签新的心肌细胞,不论其祖细胞 。 (A)的EdU掺入心肌细胞分裂过程中DNA的。增殖心肌人口将resul吨心肌的增加,或更换,因此是生产性的DNA合成(有助于组织维持和修复)。 (B)中的EdU被掺入心脏祖细胞的细胞分裂过程中的DNA。这将在细胞分化为心肌细胞谱系期间被保留。这种干细胞分化也将导致增加心肌细胞的数目,因此,有助于组织维护和修理。 (C)的心肌细胞均经过导致增加的心肌细胞的倍性,这是与心肌细胞肥大和心肌重塑相关的“非生产”DNA复制的潜能,但不替换丢失的心肌细胞。多倍体的方法,从binucleation不同,因为它导致心肌细胞与含有四个或更多组两条同源染色体(> 2N)的单核。 (D)继连续细胞核pulse,该协议描述了PCM1表达心肌细胞核的核分离和鉴定,使双方心肌套数和掺入的EdU量化。 PCM1表达,用流式细胞仪检测的EdU掺入。 请点击此处查看该图的放大版本。
Protocol
Representative Results
Discussion
要准确量化心肌营业额和再生试验必须真正心肌细胞的产生和非生产性的DNA分裂区分。许多研究继续简单地忽略这些非生产性活动,量化心肌细胞增殖仅通过周期室壁运动及细胞周期标记物的表达。迄今为止一个方法,使心肌营业额的准确定量,同时控制这些非生产性活动仍然引经据典。尤其是,它仍然很难占到心肌多倍体这有助于达到〜心肌DNA复制13的65%。因此,以协助在心肌细胞产生的精确量化,我们已经开发出一种协议,它允许新心肌细胞核的速率的健壮量化,同时排除了导致增加的倍性DNA的复制。虽然这个协议可以新的心肌细胞属间不是歧视灰和心肌细胞的双核,它可以快速地使用,并且准确地计算上限心肌代的(占倍)。因此该协议提供了一种筛选工具,以评估在疾病模型中的心肌细胞的产生和多倍体的速率潜在变化或评估治疗剂的潜在效率。一旦新心肌细胞核产生的速率的变化使用该协议确定了随后的研究可用于确定是否此由于在心肌细胞产生心肌细胞的成核数量的变化,如先前2,13,17,18所述。这些包括在脉冲期间的使用组织学定量心肌核动力学的或从的EdU脉冲动物得到在在单个核和多核心肌群体比较的EdU掺入组织切片的分析。
由于心肌成交额的低水平协议采用埃杜多次注射超过7天。这也使心肌细胞的生成所有潜在的细胞来源“追”,并允许累计心肌细胞核代量化超过这个时间段。根据研究的基础上,在此时间范围可被调整以适应心肌生成的预测水平。对的EdU掺入在心肌细胞核的准确定量,必须有与用于检测的PCM-1反应性的二级抗体的核的没有非特异性标记。因此这将是谨慎,以优化该协议的这个方面,尤其是当比其他的第二抗体建议在这个协议是用来承接附加二级抗体滴定实验。这里所描述的协议使用PCM-1的表达来识别心肌细胞核。虽然这是一个既定的心肌细胞的标记,替代标记可用于验证数据;这些包括特异性针对已被确认为心肌细胞的细胞核1部分局部心肌肌钙蛋白T抗体。同样,替代核局部蛋白可用于在比上述心肌细胞的其他核种群确定和量化的EdU掺入。重要的是,所有的心肌细胞核正在积极进行有丝分裂被排除在分析,该DNA合成的命运是未知的,并且可能导致在任一细胞分裂或增加的倍性。 PCM1是在细胞周期因此心肌经历有丝分裂不会被PCM1表达式确定的M期拆卸。此外,在细胞周期的S期的所有核应排除随后的分析。这可以通过用2N人口包括与2N和4N种群之间的DAPI强度高于DAPI强度选通所有的细胞核来实现。
虽然它是日益接受的是,心脏具有正常老化和急性损伤过程中替换心肌细胞的能力,这种潜在的源和程度仍存在争议。此外,心肌周转率不同的报道1,7,20-22。这可能部分由于在精确地确定和量化新的心肌细胞的产生19的困难。迄今为止,大多数研究仅在使用组织学分析的,并通过胞质蛋白质,包括肌节的蛋白,心肌周转和更新2,4,23,24的量化的表达心肌细胞的识别依赖。使用这些方法来检测增殖标记物的表达,或者作为这里证明的,胸苷类似物的掺入极易造成其它心脏细胞类型为心肌细胞的误认。虽然使用3D共焦成像可以帮助缓解这些问题日ESE方法是昂贵且耗时的。有趣的是,这里描述的方案展示了新的心肌细胞的核生成发生在每周0.17%的速度。这是与其它流式细胞术为基础的研究表明高达0.13%5周成交率一致。虽然这是很有诱惑力根据这些数据来推断全年周转率,因为在以往的研究2,5,25,26,作为周转率在动物13的续航时间动态,这是不合适的。
这种方法具有许多潜在的应用包括评估潜在的治疗剂,以提高心肌再生或调查心脏疾病的心肌周转和影响心肌多倍体的速率。
Divulgations
The authors have nothing to disclose.
Acknowledgements
This work was supported by the British Heart Foundation, project grant PG/13/69/30454.
Materials
| 0.32 M sucrose | Sigma | 84100 | |
| 10 mM Tris-HCl (pH = 8) | Sigma | T3253 | |
| 5 mM CaCl2 | Sigma | c5086 | |
| 5 mM magnesium acetate | sigma | M-5661 | |
| 2.0 mM EDTA | Sigma | E5134 | |
| 0.5 mM EGTA | Sigma | 63779 | |
| 1 mM DTT | Sigma | D0632 | |
| 70 mM KCl | Sigma | P9541 | |
| 10 mM MgCl2 | Sigma | M8266 | |
| 1.5 mM spermine | Sigma | 85590 | |
| Isotype rabbit IgG- ChIP Grade | abcam | abc7415 | |
| Rabbit anti-PCM-1 antibody | Sigma | HPA023374 | |
| Alexa Fluor 488 F(ab')2 Fragment of Goat Anti-Rabbit IgG (H+L) Antibody | Life technologies | A-11070 | |
| cell strainers 70 μm and 100μm | Fisher scientific | 11597522, 11517532 | |
| Glass dounce (40 ml) and pestle large clearance | Sigma | D9188-1SET | |
| EdU (5-ethynyl-2’-deoxyuridine) | Life technologies | A10044 | |
| Click-iT Plus EdU Alexa Fluor 647 Flow Cytometry Assay Kit | Life technologies | C10634 | This kit inlcudes reagents required for section, EdU reaction buffer, EdU fixative, saponin-based permeabilization solution and the reagents required for the EdU labelling cocktail. |
| CyStain DNA 2 step kit, | Sysmex Partec | 05 5005 | This kit inlcudes reagents required for DAPI labelling (DNA staining solution) |
| Probe homogeniser e.g. TissueRuptor | Qiagen | 9001273 | |
| TissueRuptor Disposable Probes | Qiagen | 990890 | |
| ultracentrifuge | Sorvall | ||
| Facscanto II | BD Biosciences | ||
| Ultracentrifuge Tube, Thinwall, Polypropylene. 38.5 mL, 25 x 89 mm | Beckman Coulter | 326823 | |
| Bovine serum albumin | Sigma | A2153 |
References
- Bergmann, O., et al. Evidence for cardiomyocyte renewal in humans. Science. 324, 98-102 (2009).
- Senyo, S. E., et al. Mammalian heart renewal by pre-existing cardiomyocytes. Nature. 493, 433-436 (2013).
- Soonpaa, M. H., Rubart, M., Field, L. J. Challenges measuring cardiomyocyte renewal. Biochim Biophys Acta. 1833, 799-803 (2013).
- Loffredo, F. S., Steinhauser, M. L., Gannon, J., Lee, R. T. Bone marrow-derived cell therapy stimulates endogenous cardiomyocyte progenitors and promotes cardiac repair. Cell Stem Cell. 8, 389-398 (2011).
- Malliaras, K., et al. Cardiomyocyte proliferation and progenitor cell recruitment underlie therapeutic regeneration after myocardial infarction in the adult mouse heart. EMBO Mol Med. 5, 191-209 (2013).
- Hsieh, P. C., et al. Evidence from a genetic fate-mapping study that stem cells refresh adult mammalian cardiomyocytes after injury. Nat Med. 13, 970-974 (2007).
- Bergmann, O., et al. Identification of cardiomyocyte nuclei and assessment of ploidy for the analysis of cell turnover. Exp Cell Res. 317, 188-194 (2011).
- Herget, G. W., Neuburger, M., Plagwitz, R., Adler, C. P. DNA content, ploidy level and number of nuclei in the human heart after myocardial infarction. Cardiovasc Res. 36, 45-51 (1997).
- Scholzen, T., Gerdes, J. The Ki-67 protein: from the known and the unknown. J Cell Physiol. 182, 311-322 (2000).
- Carmena, M., Earnshaw, W. C. The cellular geography of aurora kinases. Nat Rev Mol Cell Biol. 4, 842-854 (2003).
- Bergmann, O., Jovinge, S. Isolation of Cardiomyocyte Nuclei from Post-mortem Tissue. J. Vis. Exp. , e4205 (2012).
- Gilsbach, R., et al. Dynamic DNA methylation orchestrates cardiomyocyte development, maturation and disease. Nat Commun. 5, 5288 (2014).
- Richardson, G., Laval, S., Owens, W. A. Cardiomyocyte regeneration in the mdx mouse model of non-ischemic cardiomyopathy. Stem Cells Dev. , (2015).
- Jiao, K., et al. An essential role of Bmp4 in the atrioventricular septation of the mouse heart. Genes Dev. 17, 2362-2367 (2003).
- Bergmann, O., et al. Cardiomyocyte renewal in humans. Circ Res. 110, e17-e18 (2012).
- Prigge, J. R., et al. Nuclear double-fluorescent reporter for in vivo and ex vivo analyses of biological transitions in mouse nuclei. Mamm Genome. , (2013).
- Naqvi, N., et al. A proliferative burst during preadolescence establishes the final cardiomyocyte number. Cell. 157, 795-807 (2014).
- Liu, Z., Yue, S., Chen, X., Kubin, T., Braun, T. Regulation of cardiomyocyte polyploidy and multinucleation by CyclinG1. Circ Res. 106, 1498-1506 (2010).
- Ang, K. L., et al. Limitations of conventional approaches to identify myocyte nuclei in histologic sections of the heart. Am J Physiol Cell Physiol. 298, C1603-C1609 (2010).
- Kajstura, J., et al. Myocyte turnover in the aging human heart. Circ Res. 107, 1374-1386 (2010).
- Kajstura, J., et al. Cardiomyogenesis in the adult human heart. Circ Res. 107, 305-315 (2010).
- Walsh, S., Ponten, A., Fleischmann, B. K., Jovinge, S. Cardiomyocyte cell cycle control and growth estimation in vivo–an analysis based on cardiomyocyte nuclei. Cardiovasc Res. 86, 365-373 (2010).
- Anversa, P., Leri, A., Kajstura, J. Cardiac regeneration. J Am Coll Cardiol. 47, 1769-1776 (2006).
- Gonzalez-Valdes, I., et al. Bmi1 limits dilated cardiomyopathy and heart failure by inhibiting cardiac senescence. Nat Commun. 6, 6473 (2015).
- Kimura, W., et al. Hypoxia fate mapping identifies cycling cardiomyocytes in the adult heart. Nature. 523, 226-230 (2015).
- Soonpaa, M. H., Field, L. J. Assessment of cardiomyocyte DNA synthesis in normal and injured adult mouse hearts. Am J Physiol. 272, H220-H226 (1997).
