L’osso è un tipo di tessuto connettivo composto da due parti: organici (cellule e fibre collagene) e minerali (composti di calcio e fosfato). I componenti principali di minerali nelle ossa sono apatiti1. Diversi tipi di cellule competenti di mineralizzazione in osso (osteoblasti), nei denti (odontoblasti) e nella cartilagine (condrociti) regolano i passi iniziali di mineralizzazione di produrre proteine della matrice extracellulare (ECM) e rilasciare la matrice vescicole (MVs) (Figura 1). MVs sono 100-300 nm diametro vescicole che si accumulano di calcio e fosfato facilitando la nucleazione apatite e successivamente associare al collagene2,3. Quindi, MVs disintegrarsi per rilasciare apatiti al medium extracellulare. Le apatiti continuano a crescere a contatto con le fibre di collagene e formano la matrice dell’osso. La mineralizzazione è sostenuta dal costante rifornimento di Pio e Ca2 + nel mezzo extracellulare. Alcuni dati recentemente pubblicati supportano il nostro modello4,5. Tessuti molli non mineralizzare in condizioni fisiologiche. Tuttavia, la calcificazione ectopica può verificarsi in condizioni patologiche come calcificazione vascolare3. Le cellule vascolari che acquisiscono il fenotipo degli osteoblasti in grado di produrre MVs che inducono la nucleazione di apatiti e avviare mineralizzazione negli strati della parete mediale e intimale dei vasi sanguigni. Dal calcificazione ectopica assomigliano endochondral normale mineralizzazione3, comprendere i meccanismi molecolari della mineralizzazione delle cellule ossee e condrociti dovrebbero fornire alcuni indizi sulla calcificazione ectopica dei tessuti molli che sono formato.
Lo sviluppo di tessuti scheletrici è regolato da vari enzimi, fattori di crescita e promotori o inibitori della mineralizzazione. L’azione antagonistica di tessuto-non specifico fosfatasi alcalina (TNAP) (Figura 1) ed ectonucleotide pirofosfatasi/fosfodiesterasi ho (NPP1), insieme a ankyrin (ANK), controlla concentrazione pirofosfato inorganico (PPho) 6. PPho, un potente inibitore della formazione HA, è idrolizzato di TNAP; NPP1 idrolizza i trifosfati del nucleotide per formare PPho mentre ANK PPho Esporta dalla cella alla ECM. Il rapporto di Pi/PPi può disciplinare l’apatite formazione7,8 con possibili conseguenze patologiche9.
La membrana di MV è arricchita in proteine di trasporto dello ione che facilitano la precipitazione iniziale di calcio e fosfato all’interno la MVs durante il processo di nucleazione (Figura 1). Il trasportatore del fosfato 1 (PiT) consente di incorporare Pho generato nello spazio perivesicular nel MVs10,11. Annexins possono essere coinvolti nell’associazione e nel trasporto di Ca2 + e nel processo biofisico che avvia la mineralizzazione del MV lumen12,13. Favoriamo l’ipotesi, suggerito in precedenza, per mineralizzazione all’interno di vescicole intracitoplasmatiche di nucleazione interno di apatite dentro la MV prima della sua propagazione nell’ECM14,15. Modellazione in vitro hanno confermato l’induzione di Ca2 +/ pho complessi formazione nei proteoliposomi effettuata in PS e AnxA516. Ciò potrebbe indicare che accumulo di Ca2 +, Pho, complessi AnxA5 e PS in “lipid rafts” di membranesrepresent i microvilli-come il nucleo di nucleazione (NC) di apatite all’interno Mvs Annexins e TNAP possiedono anche collagene-associazione capacità che possono essere utili nel mettere MVs lungo le fibre di collagene e, nello stimolare la propagazione di mineralizzazione in ECM. Fetuina A e osteopontina (OPN)17, sono noti come inibitori della formazione di apatite che possono rallentare la propagazione della mineralizzazione sul patibolo collageno. Nucleazione e propagazione sono eventi distinti, il primo precede quest’ultimo, ed entrambi possono essere rilevanti per il processo di mineralizzazione patologico.
Per scoprire come la chimica dei complessi di fosfato di calcio può cambiare fisiologico mineralizzazione e calcificazione ectopica, è necessario identificare i minerali prodotti dalle cellule. Apatiti sono un gruppo di calcio e fosfato contenente minerali con cristallo generale unità cella formula Ca10(PO4) di6X2, dove X = Cl, F, OH. Essi sono classificati come segue18: fluorapatite (FA) Ca10(PO4)6F2, chlorapatite (CA) Ca10(PO4)6Cl2 e idrossiapatite (HA) Ca10(PO4 )6(OH)2.
La scelta di linee cellulari di osteoblasti per indurre la formazione di minerali è fondamentale, poiché ogni linea cellulare esibisce un profilo distinto di mineralizzazione. In questo rapporto, abbiamo confrontato la nucleazione dei minerali da due modelli di cella selezionata umana di mineralizzazione: cellule osteoblastic hFOB 1.19 e cellule di osteosarcoma Saos-2. Cellule derivate da osteosarcoma sono comunemente usate come modelli osteoblastiche e cellule Saos-2 hanno conservato il carattere osteoblastic più maturo19 mentre le cellule indifferenziate hFOB fetale umano sono ampiamente usate come un modello per il normale osteoblastic differenziazione20. I profili di mineralizzazione sono stati analizzati con metodi diversi: Alizarin Red-S (AR-S) colorazione, ultravioletta visualizzazione luce (UV), formazione immagine di microscopia elettronica (TEM) di trasmissione, quantificazione di energia dispersiva x-ray microanalisi (EDX) e dello ione mappatura. Il vantaggio di TEM-EDX su tecniche alternative utilizzate negli studi precedenti è che dà risultati quantitativi e qualitativi della sostituzione dello ione in cristalli di apatite4,5,21. L’obiettivo generale dell’utilizzo di TEM-EDX era di trovare un metodo semplice per l’imaging e quantificazione della distribuzione degli ioni Ca, F e Cl in vari minerali da diversi tipi di cellule durante fasi distinte del processo di mineralizzazione. Questo metodo è stato utilizzato con successo, ad esempio, per monitorare l’interazione delle nanoparticelle di zinco con i prodotti chimici coesistenti e loro effetti combinati su organismi acquatici22. In un altro studio, un photocatalyst rame su materiali di titanio in soluzione acquosa è stato ampiamente caratterizzato mediante spettrometria ad emissione ottica al plasma accoppiato induttivamente (ICP-OES), N2 physisorption (BET), XRD, DRS UV-vis, FT-IR, Raman spettroscopia, TEM-EDX e photoelectrochemical misure23. Il nostro obiettivo era di confrontare l’origine e la proprietà di vescicole e minerali in due linee cellulari per comprendere il meccanismo che controlla la mineralizzazione durante la differenziazione ossea.
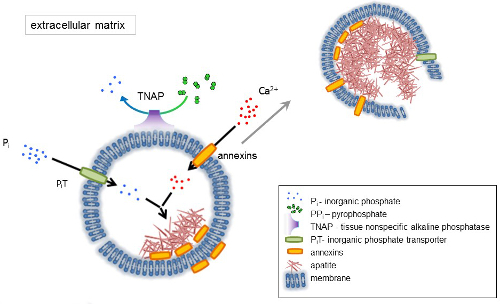
Figura 1 . Schema delle fasi iniziali di mineralizzazione in cellule ossee che coinvolge la sintesi di proteine della matrice extracellulare (ECM) ed il rilascio delle vescicole della matrice (MVs) dalla membrana. MVs accumulare calcio attraverso l’azione di proteine leganti il calcio, annexins e fosfato, attraverso l’azione di un trasportatore di fosfato inorganico (PiT) seguita dall’attività del tessuto non specifico della fosfatasi alcalina (TNAP), che defosforila PPio Pho, quindi facilitante la nucleazione apatite. Quindi, MVs si disintegrano e rilasciare apatiti al medium extracellulare. La mineralizzazione è sostenuta dal costante rifornimento di Pio e Ca2 + in extracellulare media4,5. Clicca qui per visualizzare una versione più grande di questa figura.
