Фенотипы роста вносят критический вклад в фитнес дрожжей. Естественный отбор может эффективно различать линии с темпами роста, отличающимися от обратного эффективного размера популяции, который может превышать 108 особей1. Кроме того, изменчивость темпов роста среди людей в популяции является эволюционно актуальным параметром, так как она может служить основой для стратегий выживания,таких как хеджирование ставок 2,3,4,5,6. Поэтому анализы, позволяющие проводить высокотоктяжелые измерения фенотипов роста и их распределения, имеют решающее значение для изучения микроорганизмов. Анализ роста микроколонии, описанный здесь, может генерировать индивидуальные измерения скорости роста на 105 микроколоний за эксперимент. Таким образом, этот анализ обеспечивает мощный протокол для изучения дрожжевой эволюционной генетики и геномики. Он поддается особенно хорошо для тестирования, как изменчивость в популяциях генетически идентичных одиночных клеток генерируется, поддерживается, и способствует популяции фитнес7,8,9,10.
Метод, описанный здесь (Рисунок 1) использует периодически захваченных, с низким увеличением яркое поле изображения клеток, растущих в жидких средствах массовой информации на 96- или 384-хорошо стеклянной нижней пластины для отслеживания роста в микроколонии. Клетки придерживаются лектина конканавалина А, который покрывает дно микроскопической пластины, и образуют двумерные колонии. Поскольку микроколонии растут в монослойном, область микроколонии сильно коррелирует с номером клетки7. Таким образом, точные оценки темпов роста микроколонии и времени задержки могут быть получены с помощью пользовательского программного обеспечения для анализа изображений, которое отслеживает скорость изменения площади каждой микроколонии. Кроме того, экспериментальная установка может контролировать изобилие и даже субклеточную локализацию флуоресцентно помеченных белков, выраженных в этих микроколониях. Вниз по течению обработки данных из этого микроколонии роста анализа может быть достигнуто путем пользовательского анализа или существующего программного обеспечения анализа изображений, таких как обработка изображений легко (PIE)11, алгоритм для надежного распознавания колонии области и высокой пропускной способности анализа роста с низким увеличением, яркие изображения, которая доступна через GitHub12.
Поскольку оценки темпов роста, полученные в результате анализа микроколонии роста, генерируются на основе большого числа одноколонных измерений, они чрезвычайно точны, при этом стандартные ошибки на несколько порядков меньше, чем сами оценки для эксперимента разумного размера. Таким образом, сила анализа для выявления различий в темпах роста между различными генотипами, обработками или условиями окружающей среды высока. Формат мультиэлементной пластины позволяет сравнивать множество различных комбинаций среды и генотипа в одном эксперименте. Если штаммы constitutively выражают различные флуоресцентные маркеры, они могут быть смешаны в том же хорошо и отличается последующим анализом изображения, который может увеличить мощность дальше, позволяя хорошо за колодец нормализации данных.
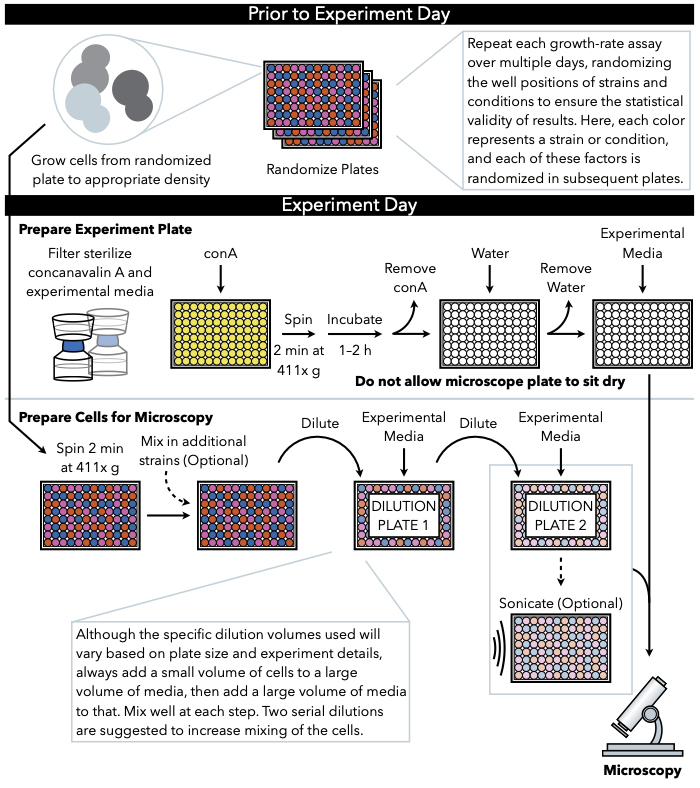
Рисунок 1: Схематическое представление протокола. Этот протокол следует за 2 главным образом шагами, которые подготовка экспериментальной плиты и подготовка клеток к изображению. Рандомизация пластин и рост клеток должны проводиться до и в течение дня эксперимента. Повторное смешивание клеток на каждом шагу во время разбавления необходимо в шагах до покрытия, и поэтому подготовка экспериментальной пластины сначала рекомендуется так, что она готова к покрытие сразу после завершения разбавления клеток. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
