반응성 산소 종(ROS) 신호는 신경계1의개발 및 기능을 조절한다. 중요한 세포 ROS 공급원은 NADPH 산화효소(NOX)로, 이는 과산화수소와 과산화수소(H2O2)2를생성하는 막 단백질이다. NOX 효소는 중추 신경계(CNS)를 통해 발견되며, NOX 유래 ROS는 뉴런 개발에기여3,4,5,6. 신경 줄기 세포의 유지 보수 및 분화, 신경 극성 확립, 중성염 아웃성장 및 시냅스 가소성은 ROS7,8,9,10,11의적절한 수준을 요구하는 것으로 나타났다. 한편, NOXes에 의한 ROS의 통제되지 않은 생산은 알츠하이머병, 다발성 경화증 및 외상성 뇌 손상을 포함한 신경 퇴행성 질환에 기여한다12,13,14. 따라서 생리학적으로 관련된 ROS의 생산은 건강한 상태를 유지하는 데 중요합니다.
유전자 인코딩 된 바이오 센서의 개발은 세포 ROS의 검출을 크게 촉진시켰습니다. 유전자 인코딩 된 바이오 센서의 한 가지 중요한 장점은 이러한 센서가 특히 뚜렷한 위치를 대상으로 할 수 있기 때문에 ROS 신호의 증가 된 시간적 및 공간 해상도입니다. 레독스에 민감한 GFP(roGFP)는 이러한 ROS 바이오 센서의 한 유형입니다. roGFP2-Orp1 변이체는 효모15,16에서글루타티온 peroxiredoxin 가족 단백질인 Orp1 도메인을 통해 H2O2를 구체적으로 검출한다. Orp1 단백질의 산화는 그것의 형성을 바꾸기 위하여 roGFP2로 옮겨져(도 1A). 이 프로브는 405nm와 480nm 근처의 두 개의 여기 봉우리와 515 nm에서 단일 방출 피크를 전시합니다. 산화 시, 흥분 피크 주위 형광 강도 변경: 동안 405 nm 흥분 증가, 480 nm 흥분 감소. 따라서, roGFP2-Orp1은 비율인식 생체센서이며,H2O-2-레벨은두 개의 상이한 파장(도1B)에서흥분되는 형광 강도의 비율에 의해 검출된다. 전반적으로 roGFP2-Orp1은 생체 내에서효율적으로 활용할 수 있는 ROS 이미징을 위한 다목적 도구입니다.
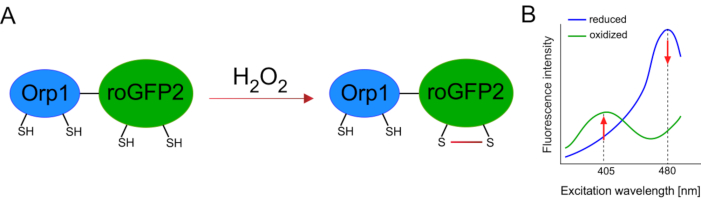
그림 1: roGFP2-Orp1의 회로도 표현 및 여기 스펙트럼. (A)Oxidant 전송은 H 2 O2에대한 응답으로 Orp1과 roGFP2 사이에 발생하여 roGFP2의 형성적 변화를 초래한다. (B)roGFP2-Orp1의 흥분 스펙트럼은 405 nm와 480 nm및 515 nm에서 단일 방출 피크에서 두 개의 여기 피크를 나타낸다. H2O2에의해 산화시, 405 nm 흥분 증가 하는 동안 480 nm 흥분 감소. 이렇게 하면 H2 O2존재에 대한 비율 측정 판독이 발생합니다. 이 수치는 빌란과 벨로소프(2017)16및 모건 외(2011)25에서수정되었다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
Danio rerio (제브라피시) 모델 시스템은 유전자 인코딩 된 바이오 센서를 적용하는 데 몇 가지 장점이 있습니다. 배아와 애벌레의 광학적 투명성은 비침습적 생체 내 이미징을 가능하게 합니다. 새로운 이미징 도구는 더 높은 해상도와 더 깊은 침투를 달성하기 위해 개발되고있다17. 또한, 유전자 조작을 위한 확립된 도구(ectopic mRNA 발현, 톨2 전세대 등) 및 게놈 편집(TALENs, CRISPR/Cas9 등) 및 게놈 편집(TALEN, CRISPR/Cas9 등)이 있어 트랜스제닉 동물의 생성을 촉진하는18. 제브라피쉬 배아가 어머니 밖에서 발전함에 따라 이 시스템은 배아의 접근과 조작을 더욱 쉽게 할 수 있습니다. 예를 들어, 1 세포 단계 주사 및 약물 치료는 쉽게 할 수 있습니다.
여기서, 우리는 체외 전사 mRNA에 주입하여 H2O2-특정바이오 센서 roGFP2-Orp1을 일시적으로 표현하기 위해 제브라피쉬를 사용했습니다. 이러한 배아는 배양된 뉴런의 체외 이미징과 생체 내이미징(도 2)에 모두 사용될 수 있다. 우리는 제브라피시 배아에서 망막 신경절 세포 (RGC)를 해부하고 도금하기위한 프로토콜을 설명하고 배양 뉴런에서 H2O2-수준을 평가합니다. 그런 다음, 우리는 공초점 현미경 검사를 사용하여 roGFP2-Orp1 표현 배아 및 애벌레의 생체 내 이미징을 위한 방법을 제시합니다. 이 접근은 생리적인 H2O2-수준을 결정할 뿐만 아니라 다른 발달 단계 또는 조건에서 생기는 잠재적인 변경을 결정할 수 있습니다. 전반적으로, 이 시스템은 살아있는 세포및 동물에서H2O2를 검출하는 신뢰할 수 있는 방법을 제공하여 개발, 건강 및 질병에서 H 2 O2의역할을 연구한다.
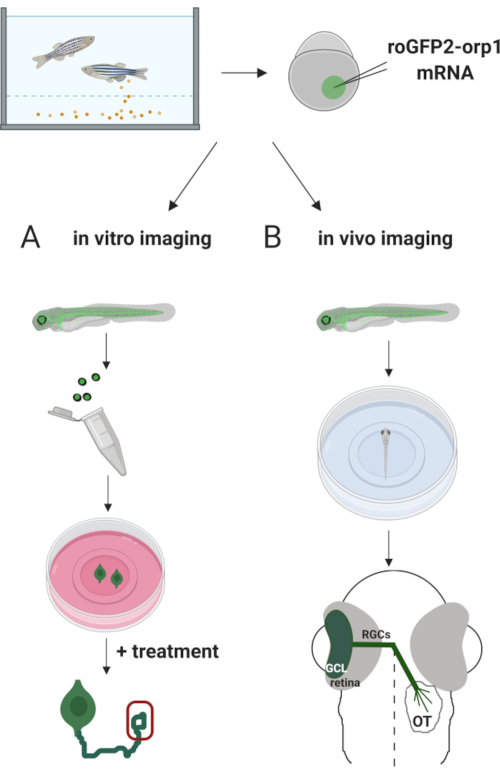
그림 2. 실험적 접근 방식의 개요입니다. 간단히, 배아 수집 후, roGFP2-Orp1 mRNA는 1 세포 단계 제브라피시 배아의 노른자로 주입된다. 개발 배아는 생체 외에서 (A)및(B)생체 내 이미징 모두에 사용할 수 있습니다. (A)GFP 양성 배아는 34hpf에서 RGC 수집을 위한 망막을 해부하는 데 사용된다. 해리된 RG는 ZFCM(+) 미디어의 PDL/라미닌 코팅 커버립에 도금됩니다. RGCs가 도금 6-24 h 후 축축을 확장함에 따라 성장 콘 이미징을 수행할 수 있습니다. 세포는 H2O 2-level의 잠재적인 변화를 측정하기 위해 다른 치료를 받을 수 있습니다. 여기서, 우리는 RGC (빨간색)의 성장 콘에서 H2O2-수준을 측정했습니다. (B)GFP 양성 배아는 생체 내 이미징에 사용된다. 원하는 나이에, 태아는 공초점 화상 진찰을 위한 35 mm 유리 바닥 접시에 마취하고 장착될 수 있습니다. 여기서 배아는 망막 이미징을 위해 통풍구로 장착됩니다. 회로도는 제브라피시의 망막 발달을 보여줍니다. RGC는 망막에서 가장 안쪽 층인 신경절 세포 층(GCL)을 형성합니다. RGC 축축은 시신경으로 발전하여 중선을 넘어 시구를 형성합니다. 그런 다음 RGC 축축은 중뇌의 광학 텍텀에서 시냅스를 만들기 위해 등등으로 자랍니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
