活性酸素種(ROS)シグナル伝達は神経系1の発達および機能を調節する。重要な細胞ROS源は、過酸化水素と過酸化水素を生成する膜貫通タンパク質であるNADPHオキシダーゼ(NOX)である(H2O2)2。NOX酵素は中枢神経系(CNS)全体に見られ、NOX由来のROSは神経細胞の発達に寄与する3、4、5、6。神経幹細胞の維持と分化、神経極性の確立、神経突起伸長、およびシナプス可塑性は、ROS7、8、9、10、11の十分なレベルを必要とすることが示されている。一方、NOXesによるROSの制御不能な産生は、アルツハイマー病、多発性硬化症、および外傷性脳損傷12、13、14を含む神経変性疾患に寄与する。したがって、生理学的に関連するROSの産生は、健康な状態を維持するために重要である。
遺伝子組み換えバイオセンサーの開発は、細胞ROSの検出を大いに促進した。遺伝子組み換えバイオセンサーの重要な利点の1つは、ROS信号の時間的および空間分解能の増加であり、これらのセンサーは特異的に異なる場所を標的とすることができる。レドックス感受性GFP(roGFP)は、ROSバイオセンサーのような一種である。roGFP2-Orp1変異体は、特にそのOrp1ドメインを介してH2O2を検出し、これは酵母15、16由来のグルタチオンペルオキシレドキシンファミリータンパク質である。Orp1タンパク質の酸化は、その立体構造を変化させるためにroGFP2に移される(図1A)。このプローブは、405 nmおよび480 nm付近の2つの励起ピークと、515 nmの単一の放出ピークを示す。酸化すると、励起ピーク周辺の蛍光強度が変化する:405nm励起が増加する一方で、480nmの励起は減少する。従って、roGFP2-Orp1は、レシオメトリックバイオセンサであり、H2O2-レベルは、2つの異なる波長で励起された蛍光強度の比によって検出される(図1B)。全体的に見て、roGFP2-Orp1は、インビボで効率的に利用することができるROSイメージング用の汎用性の高いツールです。
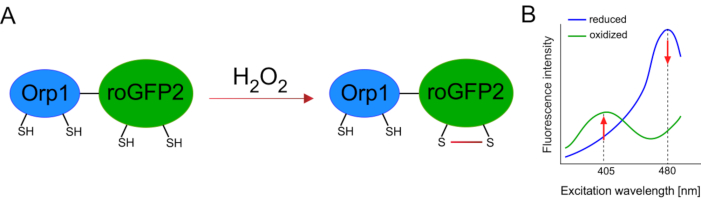
図1: roGFP2-Orp1の模式的表現と励起スペクトル(A)は、H2O2に応答してOrp1とroGFP2の間で酸化伝達が起こり、roGFP2の立体構造変化をもたらす。(B)roGFP2-Orp1の励起スペクトルは、405 nmおよび480 nmの2つの励起ピークと515 nmの単一放出ピークを示す。H2O2による酸化の際、405nm励起は増加し、480nm励起は減少する。この結果、H2 O2の比率の読み出しが得られます。この数字は、ビランとベロウソフ(2017)16とモルガンら(2011)25から変更されています。この図の大きなバージョンを表示するには、ここをクリックしてください。
Danio rerio(ゼブラフィッシュ)モデルシステムは、遺伝的にコード化されたバイオセンサーを適用するためのいくつかの利点を有する。胚および幼虫の光学的透明性は、生体内での非侵襲的なイメージングを可能にする。新しいイメージングツールは、より高い解像度と深い浸透を達成するために開発されています17.さらに、遺伝子操作(異所性mRNA発現、Tol2トランスジェネシス等)やゲノム編集(タレンス、CRISPR/Cas9等)の樹立されたツールがあり、トランスジェニック動物18の生成を促進する。ゼブラフィッシュの胚が母親の外で発達するにつれて、このシステムはさらに胚の容易なアクセスおよび操作を可能にする。例えば、1細胞段階の注射や薬物治療は簡単に行うことができます。
ここでは、ゼブラフィッシュを用いて、生体転写mRNAを注入してH2O2特異的バイオセンサーroGFP2-Orp1を一時的に発現させた。これらの胚は、培養ニューロンのインビトロイメージングとインビボイメージングの両方に使用することができる(図2)。ゼブラフィッシュ胚からの解剖およびメッキのレチナル神経節細胞(RGC)のプロトコルを説明し、続いて培養ニューロンにおけるH2O2-レベルを評価する。次に、共焦点顕微鏡を用いてroGFP2-Orp1発現胚および幼虫のインビボイメージング法を提示する。このアプローチは、生理学的なH2O2-レベルを決定するだけでなく、異なる発達段階または条件で起こる潜在的な変化を決定することを可能にする。全体として、このシステムは、生きた細胞および動物におけるH2O2を検出するための信頼できる方法を提供し、H2O2の発達、健康および疾患における役割を研究する。
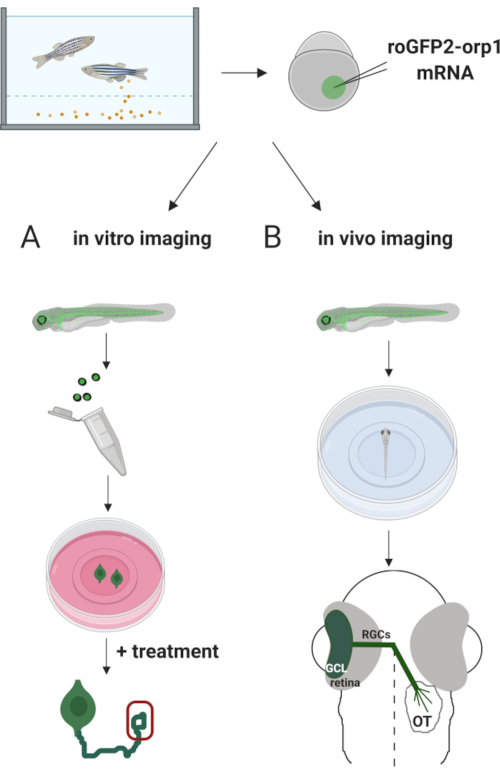
図 2.実験アプローチの概要簡単に言えば、胚採取後、roGFP2-Orp1 mRNAを、ゼブラフィッシュ胚の1細胞期の黄身に注入する。現像胚は、インビトロ(A)および(B)インビボイメージングの両方に使用することができる。(A) GFP陽性胚は、34 hpfでのRGC採取のためのレティナを解剖するために使用される。解約されたRGCは、ZFCM(+)メディアのPDL/ラミニンコーティングカバーリップにメッキされています。成長コーンイメージングは、GMCが6〜24時間のめっき後に軸索を延長するにつれて行うことができる。細胞は、H2O2-レベルにおける潜在的変化を測定するために異なる治療を施すことができる。ここでは、RGCの成長コーンにおけるH2O2-レベル(赤色)を測定した。(B)GFP陽性胚は生体内イメージングに使用される。望ましい年齢で、胚は共焦点のイメージ投射のための35のmmガラスの底の皿に麻酔され、取付けることができる。ここでは、胚は、画像の再生のために腹孔に取り付けられている。概略図はゼブラフィッシュのレチナルの発達を示す。GGCは、残膜の最も内側の層であるガングリオン細胞層(GCL)を形成する。RGC軸索は、正中線を横切るために視神経に発達し、視神経を形成する。その後、RGC軸索は、中脳の視状体でシナプスを作るためにドーサリーに成長する。この図の大きなバージョンを表示するには、ここをクリックしてください。
