أنواع الأكسجين التفاعلية (ROS) الإشارات ينظم تطوير وأداء الجهاز العصبي1. مصدر ROS الخلوية الهامة هي NADPH oxidases (NOX)، والتي هي بروتينات ترانسميمبرين توليد سوبر أكسيد وبيروكسيد الهيدروجين (H2O2)2. توجد إنزيمات NOX في جميع أنحاء الجهاز العصبي المركزي (CNS) ، ويساهم ROS المشتقة من NOX في تطوير الخلايا العصبية3و4و5و6. وقد ثبت صيانة وتمايز الخلايا الجذعية العصبية، وإنشاء قطبية الخلايا العصبية، و نمو نيوريت، و اللدونة متشابك تتطلب مستويات كافية من ROS7،8،9،10،11. من ناحية أخرى، إنتاج غير المنضبط من ROS بواسطة NOXes تسهم في الاضطرابات العصبية بما في ذلك مرض الزهايمر، والتصلب المتعدد، وإصابات الدماغالرضية 12،13،14. وبالتالي، فإن إنتاج ROS ذات الصلة من الناحية الفسيولوجية أمر بالغ الأهمية للحفاظ على ظروف صحية.
وقد سهل تطوير أجهزة الاستشعار البيولوجية المشفرة وراثيا الكشف عن ال ROS الخلوية بشكل كبير. ومن المزايا الهامة لأجهزة الاستشعار البيولوجية المشفرة وراثيا زيادة الاستبانة الزمنية والمكانية لإشارة ROS، حيث يمكن استهداف أجهزة الاستشعار هذه على وجه التحديد إلى مواقع متميزة. GFP الحساسة للأكسدة (roGFP) هو نوع واحد من هذه أجهزة الاستشعار الحيوية ROS. البديل roGFP2-Orp1 يكشف على وجه التحديد H2O2 من خلال مجالها Orp1، وهو بروتين الأسرة بيروكسيريدوكسيني الجلوتاثيون من الخميرة15،16. يتم نقل أكسدة البروتين Orp1 إلى roGFP2 لتغيير تشكيله (الشكل 1A). ويعرض المسبار قمتين للإثارة بالقرب من 405 نانومتر و480 نانومتر، وذروة انبعاث واحدة عند 515 نانومتر. عند الأكسدة ، تتغير كثافة الفلورسينس حول قمم الإثارة: في حين يزيد الإثارة 405 نانومتر ، ينخفض 480 نانومتر من الإثارة. وهكذا، roGFP2-Orp1 هو حساس حيوي نسبة، ويتم الكشف عن H2O2-levels بنسبة كثافة الفلورية متحمس في أطوال موجية مختلفة اثنين(الشكل 1B). عموما، roGFP2-Orp1 هو أداة متعددة الاستخدامات للتصوير ROS التي يمكن استخدامها بكفاءة في الجسم الحي.
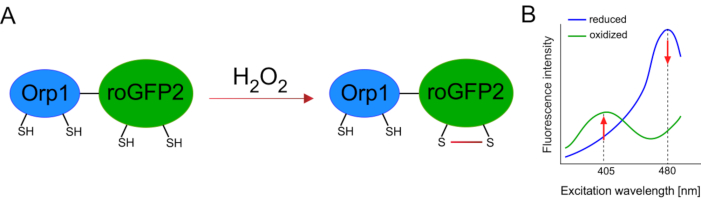
الشكل 1: التمثيل التخطيطي وأطياف الإثارة من roGFP2-Orp1. (A) نقل الأكسدة يحدث بين Orp1 و roGFP2 ردا على H2O2, مما يؤدي إلى تغييرات تشكيلية في roGFP2. (ب) تعرض أطياف الإثارة من roGFP2-Orp1 قمتين للإثارة عند 405 نانومتر و480 نانومتر وذروة انبعاث واحدة عند 515 نانومتر. عند الأكسدة بواسطة H2O2، يزيد الإثارة 405 نانومتر في حين ينخفض 480 نانومتر من الإثارة. ينتج عن ذلك قراءات معدلية لوجود H2O2. تم تعديل الرقم من بيلان وبيلوسوف (2017)16 ومورغان وآخرين (2011)25. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
نظام نموذج دانيو ريو (حمار وحشي) له العديد من المزايا لتطبيق أجهزة الاستشعار البيولوجية المشفرة وراثيا. الشفافية البصرية للأجنة واليرقات تمكن التصوير غير الغازي في الجسم الحي. ويجري تطوير أدوات التصوير الجديدة لتحقيق دقة أعلى وأعمق اختراق17. وعلاوة على ذلك، هناك أدوات راسخة للتلاعب الجيني (التعبير ميرنا خارج الرحم، Tol2 transgenesis، الخ) وتحرير الجينوم (TALENs، CRISPR/Cas9، الخ)، الذي يعزز جيل الحيوانات المعدلةوراثيا 18. كما تتطور أجنة حمار وحشي خارج الأم، وهذا النظام يسمح كذلك سهولة الوصول والتلاعب في الأجنة. على سبيل المثال، يمكن بسهولة حقن المرحلة من خلية واحدة والعلاجات الدوائية.
هنا، استخدمنا حمار وحشي للتعبير عن عابر H2O2-محددة الاستشعار الحيوي roGFP2-Orp1 عن طريق حقن في الحمض النووي الريبي المختبرة المنسوخة. ويمكن استخدام هذه الأجنة في كل من التصوير المختبري للخلايا العصبية المستزرعة والتصوير في الجسم الحي (الشكل 2). نحن نصف بروتوكولا لتشريح وصفيح خلايا العقدة الشبكية (RGCs) من أجنة حمار وحشي يليه تقييم H2O2-levels في الخلايا العصبية المستزرعة. ثم نقدم طريقة للتصوير الحي للأجنة واليرقات المعبرة عن roGFP2-Orp1 باستخدام المجهر البؤري. هذا النهج لا يسمح فقط لتحديد الفسيولوجية H2O2-مستويات ولكن أيضا التغيرات المحتملة التي تحدث في مراحل النمو المختلفة أو الظروف. بشكل عام، يوفر هذا النظام طريقة موثوقة للكشف عن H2O2 في الخلايا الحية والحيوانات لدراسة دور H2O2 في التنمية والصحة والمرض.
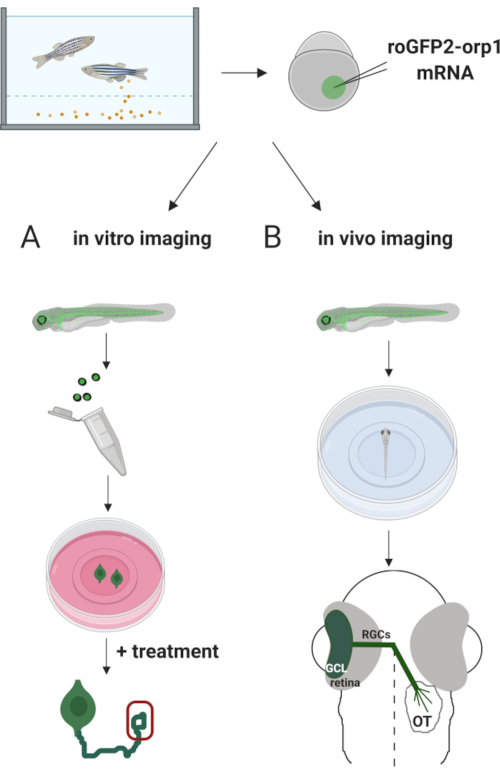
الشكل 2. الخطوط العريضة للنهج التجريبي. لفترة وجيزة، بعد جمع الأجنة، يتم حقن roGFP2-Orp1 مرنا في صفار من خلية واحدة مرحلة أجنة حمار وحشي. يمكن استخدام الأجنة النامية لكل من (أ) في المختبر و (ب) في التصوير الحي. (أ)تستخدم الأجنة الإيجابية GFP لتشريح شبكية العين لجمع RGC في 34 hpf. لوحات RGCs المفككة على أغطية مغلفة ب PDL/laminin في وسائط ZFCM (+). يمكن إجراء تصوير مخروط النمو حيث تقوم RGCs بتوسيع محاورها المحورية بعد 6-24 ساعة من الطلاء. يمكن أن تخضع الخلايا لعلاجات مختلفة لقياس التغيرات المحتملة في H2O2-levels. هنا، قمنا بقياس H2O2-مستويات في المخاريط نمو RGCs (الأحمر). (ب)تستخدم الأجنة الإيجابية GFP للتصوير في الجسم الحي. في العمر المطلوب، يمكن تخدير الأجنة وتركيبها على أطباق قاع زجاجية مقاس 35 ملم للتصوير البؤري. هنا، يتم تركيب الأجنة بطنية لتصوير الشبكية. التخطيطي يظهر تطور الشبكية في حمار وحشي. تشكل RGCs طبقة خلايا العقدة (GCL) ، وهي الطبقة الأعمق في شبكية العين. محاور RGC تتطور إلى العصب البصري لعبور خط الوسط، وتشكيل chiasm البصرية. ثم، محاور RGC تنمو ظهريا لجعل نقاط الاشتباك العصبي في استئصال البصرية في الدماغ المتوسط. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
