Reaktive iltarter (ROS) signalering regulerer udviklingen og funktionen af nervesystemet1. En vigtig cellulær ROS-kilde er NADPH-oxider (NOX), som er transmembranproteiner, der genererer superoxid og hydrogenperoxid (H2O2)2. NOX enzymer findes i hele centralnervesystemet (CNS), og NOX-afledt ROS bidrager til neuronal udvikling3,4,5,6. Vedligeholdelse og differentiering af neurale stamceller, etablering af neuronal polaritet, neuritvækst og synaptisk plasticitet har vist sig at kræve passende niveauer af ROS7,8,9,10,11. På den anden side bidrager ukontrolleret produktion af ROS af NOX’er til neurodegenerative lidelser, herunder Alzheimers sygdom, multipel sklerose og traumatisk hjerneskade12,13,14. Derfor er produktion af fysiologisk relevant ROS afgørende for at opretholde sunde forhold.
Udvikling af genetisk kodede biosensorer lettede detektionen af cellulær ROS meget. En vigtig fordel ved genetisk kodede biosensorer er den øgede tidsmæssige og rumlige opløsning af ROS-signalet, da disse sensorer specifikt kan målrettes mod forskellige steder. Redox-følsomme GFP (roGFP) er en type af sådanne ROS biosensorer. Den roGFP2-Orp1 variant specifikt registrerer H2O2 gennem sin Orp1 domæne, som er en glutathion peroxiredoxin familie protein fra gær15,16. Oxidationen af Orp1-proteinet overføres til roGFP2 for at ændre dets kropsbygning (Figur 1A). Sonden udviser to excitation toppe nær 405 nm og 480 nm, og en enkelt emission peak på 515 nm. Ved oxidation ændres fluorescensintensiteten omkring excitationstoppene: mens 405 nm excitation øges, falder 480 nm excitation. RoGFP2-Orp1 er således en ratiometrisk biosensor, og H2O2-niveaueropdages ved forholdet mellem fluorescensintensiteter, der er ophidset ved to forskellige bølgelængder (figur 1B). Samlet set roGFP2-Orp1 er et alsidigt værktøj til ROS billedbehandling, der kan udnyttes effektivt in vivo.
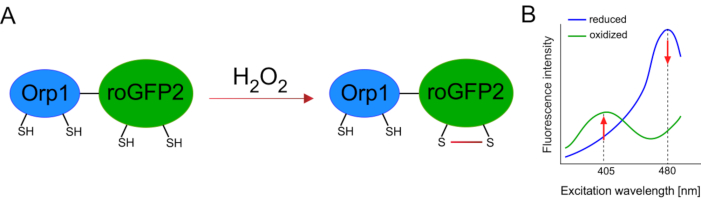
Figur 1: Skematisk repræsentation og excitationsspektre af roGFP2-Orp1. (A) Oxidantoverførsel finder sted mellem Orp1 og roGFP2 som svar på H2O2, hvilket fører til konformationsmæssige ændringer i roGFP2. (B) Den excitation spektre af roGFP2-Orp1 udviser to excitation toppe på 405 nm og 480 nm og enkelt emission peak på 515 nm. Ved oxidation med H2O2øges 405 nm excitationen, mens 480 nm excitation falder. Dette resulterer i en ratiometrisk udlæsning for H2O2 tilstedeværelse. Tallet er blevet ændret fra Bilan og Belousov (2017)16 og Morgan et al. (2011)25. Klik her for at se en større version af dette tal.
Danio rerio (zebrafisk) modelsystemet har flere fordele ved at anvende genetisk kodede biosensorer. Den optiske gennemsigtighed af embryoner og larver muliggør ikke-invasiv in vivo-billeddannelse. Der udvikles nye billedbehandlingsværktøjer for at opnå højere opløsning og dybere gennemtrængning17. Desuden findes der etablerede værktøjer til genmanipulation (ektopisk mRNA-ekspression, Tol2-transgenese osv.) og genomredigering (TALEN’er, CRISPR/Cas9 osv.), som fremmer generering af transgene dyr18. Efterhånden som zebrafiskembryoner udvikler sig uden for moderen, giver dette system yderligere lettere adgang til og manipulation af embryonerne. For eksempel kan en-celle fase injektioner og medicinske behandlinger nemt gøres.
Her brugte vi zebrafisk til forbigående at udtrykke H2O2-specifikbiosensor roGFP2-Orp1 ved at injicere in vitro-transskriberet mRNA. Disse embryoner kan anvendes til både in vitro-billeddannelse af dyrkede neuroner og in vivo-billeddannelse (figur 2). Vi beskriver en protokol til dissekering og plating nethinde ganglion celler (RGCs) fra zebrafisk embryoner efterfulgt af en vurdering H2O2-niveaueri dyrkede neuroner. Derefter præsenterer vi en metode til in vivo-billeddannelse af roGFP2-Orp1-udtrykkende embryoner og larver ved hjælp af konfokal mikroskopi. Denne tilgang gør det ikke kun muligt at bestemme fysiologiske H2O2-niveauer,men også potentielle ændringer, der forekommer i forskellige udviklingsstadier eller forhold. Samlet set giver dette system en pålidelig metode til påvisning af H2O2 i levende celler og dyr til at studere H 2 O2’srolle inden for udvikling, sundhed og sygdom.
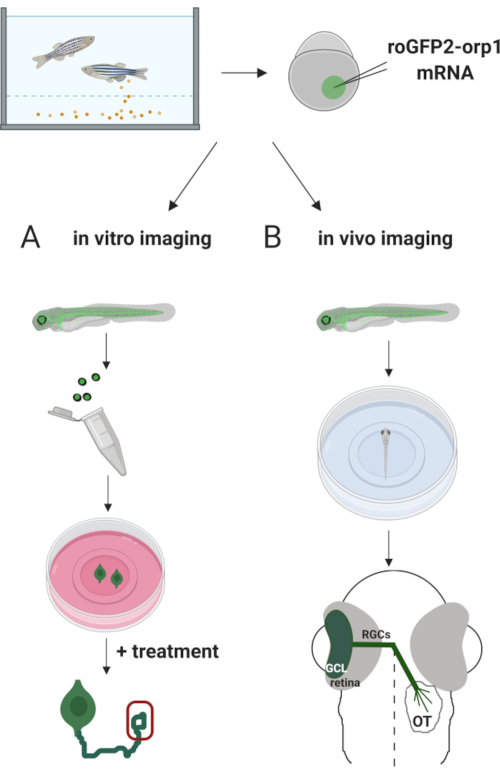
Figur 2. Oversigt over den eksperimentelle tilgang. Kort efter embryonindsamling injiceres roGFP2-Orp1 mRNA i æggeblommen på zebrafiskembryoner i en cellefase. Udvikling af embryoner kan anvendes til både (A) in vitro og (B) in vivo billeddannelse. (A) GFP-positive embryoner anvendes til dissekere nethinder til RGC-indsamling ved 34 hkf. Dissociated RGCs er belagt på PDL/laminin-coated coverslips i ZFCM (+) medier. Vækst kegle billeddannelse kan udføres som RGCs udvide deres axoner efter 6-24 timer af plating. Celler kan udsættes for forskellige behandlinger for at måle de potentielle ændringer i H2O2-niveauer. Her målte vi H2O2-niveauer i vækstkeglerne på RDC’er (rød). (B) GFP-positive embryoner anvendes til in vivo-billeddannelse. I den ønskede alder kan embryoner bedøves og monteres på 35 mm glasbundretter til konfokal billeddannelse. Her er embryoner monteret ventrally for nethinde billeddannelse. Skematisk viser nethinde udvikling i zebrafisk. RGCs danner ganglion cellelag (GCL), som er det inderste lag i nethinden. RGC axoner udvikler sig til synsnerve til at krydse midterlinjen og danner optisk chiasm. Derefter vokser RGC axoner dorsalt for at lave synapser i det optiske tectum i mellemskraben. Klik her for at se en større version af dette tal.
