La signalisation réactive des espèces de l’oxygène (ROS) régule le développement et le fonctionnement du système nerveux1. Une source importante de ROS cellulaires sont les oxydases NADPH (NOX), quisont des protéines transmembranaires générant du superoxyde et du peroxyde d’hydrogène (H2O2)2. Les enzymes NOX se trouvent dans tout le système nerveux central (SNC), et les ROS dérivés du NOX contribuent au développement neuronal3,4,5,6. Il a été démontré que le maintien et la différenciation des cellules souches neurales, l’établissement de la polarité neuronale, l’excroissance des neurites et la plasticité synaptique nécessitent des niveaux adéquats de ROS7,8,9,10,11. D’autre part, la production incontrôlée de ROS par les NEX contribue aux troubles neurodégénératifs, y compris la maladie d’Alzheimer, la sclérose en plaques et les lésions cérébrales traumatiques12,13,14. Par conséquent, la production de ROS physiologiquement pertinents est essentielle au maintien de conditions saines.
La mise au point de biocapteurs génétiquement codés a grandement facilité la détection des ROS cellulaires. Un avantage important des biocapteurs génétiquement codés est l’augmentation de la résolution temporelle et spatiale du signal ROS, car ces capteurs peuvent être spécifiquement ciblés à des endroits distincts. Le GFP sensible aux redox (roGFP) est l’un des types de ces biocapteurs ROS. La variante roGFP2-Orp1 détecte spécifiquement H2O2à travers son domaine Orp1, qui est une protéine de la famille des glutathion peroxiredoxin de levure15,16. L’oxydation de la protéine Orp1 est transférée dans roGFP2 pour modifier sa conformation(figure 1A). La sonde présente deux pics d’excitation proches de 405 nm et 480 nm, et un seul pic d’émission à 515 nm. Lors de l’oxydation, l’intensité de fluorescence autour des pics d’excitation change: alors que l’excitation de 405 nm augmente, l’excitation de 480 nm diminue. Ainsi, roGFP2-Orp1 est un biocapteur ratiométrique,et les niveauxH2O2sont détectés par le rapport des intensités de fluorescence excitées à deux longueurs d’onde différentes(figure 1B). Dans l’ensemble, roGFP2-Orp1 est un outil polyvalent pour l’imagerie ROS qui peut être utilisé efficacement in vivo.
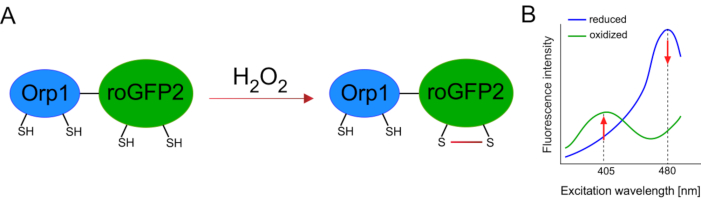
Figure 1: Spectres schématiques de représentation et d’excitation de roGFP2-Orp1. (A) Le transfert d’oxydant se produit entre Orp1 et roGFP2 en réponse àH2O2,conduisant à des changements conformationnels dans roGFP2. (B) Les spectres d’excitation du roGFP2-Orp1 présentent deux pics d’excitation à 405 nm et 480 nm et un pic d’émission unique à 515 nm. Lors de l’oxydation parH2O2,l’excitation de 405 nm augmente tandis que l’excitation de 480 nm diminue. Il en résulte une lecture ratiométrique pour la présenceH2O2. Le chiffre a été modifié à partir de Bilan et Belousov (2017)16 et Morgan et coll. (2011)25. Veuillez cliquer ici pour voir une version plus grande de cette figure.
Le système modèle Danio rerio (poisson zèbre) présente plusieurs avantages pour l’application de biocapteurs génétiquement codés. La transparence optique des embryons et des larves permet une imagerie in vivo non invasive. De nouveaux outils d’imagerie sont en cours de développement pour atteindre une résolution plus élevée et une pénétration plus profonde17. En outre, il existe des outils établis pour la manipulation génétique (expression ectopique de l’ARNm, transgénèse Tol2, etc.) et l’édition du génome (TALENs, CRISPR/Cas9, etc.), qui favorisent la génération d’animaux transgéniques18. Au fur et à mesure que les embryons de poisson zèbre se développent à l’extérieur de la mère, ce système facilite l’accès et la manipulation des embryons. Par exemple, les injections au stade unicellulaire et les traitements médicamenteux peuvent facilement être effectués.
Ici, nous avons utilisé le poissonzèbre pour exprimer transitoirement le biocapteur roGFP2-Orp1 spécifique àH2O2en injectant de l’ARNm transcrit in vitro. Ces embryons peuvent être utilisés à la fois pour l’imagerie in vitro de neurones cultivés et pour l’imagerie in vivo (figure 2). Nous décrivons un protocole pour disséquer et plaquer les cellules rétiniennes de ganglion (RGCs) des embryons de poisson zèbre suivis d’évaluer H2O2– niveaux dans les neurones cultivés. Ensuite, nous présentons une méthode d’imagerie in vivo d’embryons et de larves exprimant roGFP2-Orp1 à l’aide de la microscopie confocale. Cette approche permet non seulement de déterminer les niveaux physiologiques deH2O2-mais aussi les changements potentiels survenant à différents stades ou conditions de développement. Dans l’ensemble, ce système fournit une méthode fiable pour détecterH2O2dans les cellules vivantes et les animaux pour étudier le rôle de H2O2 dans le développement, la santé et la maladie.
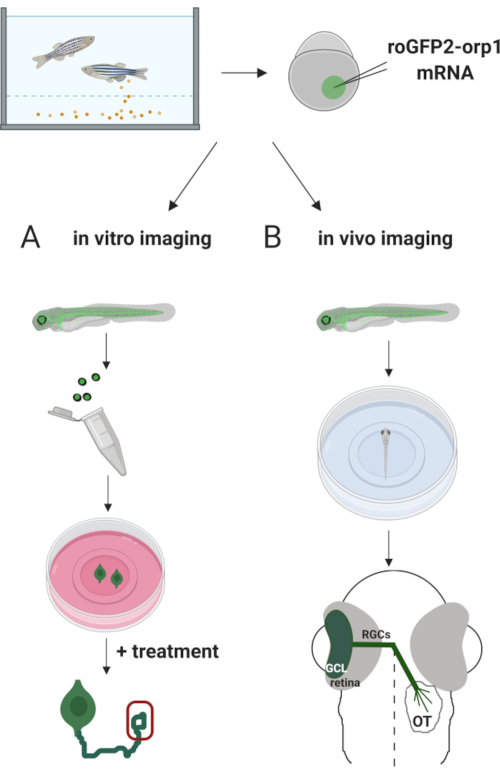
Figure 2. Aperçu de l’approche expérimentale. En bref, après le prélèvement d’embryons, l’ARNm roGFP2-Orp1 est injecté dans le jaune d’embryons de poisson zèbre au stade cellulaire. Les embryons en développement peuvent être utilisés à la fois pour l’imagerie in vitro (A)et(B) in vivo. (A) Les embryons GFP-positifs sont utilisés pour disséquer des rétines pour la collecte de RGC à 34 hpf. Les RGCs dissociés sont plaqués sur des lamaux de couverture enduits de PDL/laminine dans des milieux ZFCM (+). L’imagerie du cône de croissance peut être effectuée car les RGCs étendent leurs axones après 6-24 h de placage. Les cellules peuvent être soumises à différents traitements pour mesurer les changements potentiels des niveaux deH2O2. Ici, nous avons mesuréles niveaux deH2O2dans les cônes de croissance des RGCs (rouge). (B) les embryons positifs à la GFP sont utilisés pour l’imagerie in vivo. À l’âge souhaité, les embryons peuvent être anesthésiés et montés sur des plats à fond de verre de 35 mm pour l’imagerie confocale. Ici, les embryons sont montés ventralement pour l’imagerie rétinienne. Le schéma montre le développement rétinien chez le poisson zèbre. Les RGCs forment la couche cellulaire ganglionnaire (GCL), qui est la couche la plus interne de la rétine. Les axones de RGC se développent en nerf optique pour croiser le midline, formant le chiasm optique. Ensuite, les axones RGC se développent dorsalement pour faire des synapses dans le tectum optique dans le mécérine. Veuillez cliquer ici pour voir une version plus grande de cette figure.
