Reaktive oksygenarter (ROS) som signaliserer regulerer utvikling og funksjon av nervesystemet1. En viktig cellulær ROS-kilde er NADPH oksidase (NOX), som er transmembrane proteiner som genererer superoksid og hydrogenperoksid(H2O2)2. NOX enzymer finnes i hele sentralnervesystemet (CNS), og NOX-avledet ROS bidrar til nevronutvikling3,4,5,6. Vedlikehold og differensiering av nevrale stamceller, etablering av nevronal polaritet, nevrittutvekst og synaptisk plastisitet har vist seg å kreve tilstrekkelige nivåer av ROS7,8,9,10,11. På den annen side bidrar ukontrollert produksjon av ROS av NOXer til nevrodegenerative lidelser, inkludert Alzheimers sykdom, multippel sklerose og traumatisk hjerneskade12,13,14. Derfor er produksjon av fysiologisk relevant ROS avgjørende for å opprettholde sunne forhold.
Utvikling av genetisk kodede biosensorer forenklet påvisning av cellulær ROS sterkt. En viktig fordel med genetisk kodede biosensorer er den økte tidsmessige og romlige oppløsningen til ROS-signalet, da disse sensorene kan målrettes spesifikt mot forskjellige steder. Redoksensitiv GFP (roGFP) er en type slike ROS biosensorer. RoGFP2-Orp1-varianten oppdager spesielt H2O2 gjennom Orp1-domenet, som er et glutathione peroxiredoxin-familieprotein fra gjær15,16. Oksidasjonen av Orp1-proteinet overføres til roGFP2 for å endre konformasjonen (figur 1A). Sonden viser to eksitasjonstopper nær 405 nm og 480 nm, og en enkelt utslippstopp på 515 nm. Ved oksidasjon endres fluorescensintensiteten rundt eksitasjonstopper: mens 405 nm eksitasjon øker, reduseres 480 nm eksitasjon. Dermed er roGFP2-Orp1 en ratiometrisk biosensor, og H2O2-nivåer oppdages av forholdet mellom fluorescensintensiteter begeistret ved to forskjellige bølgelengder (Figur 1B). Totalt sett er roGFP2-Orp1 et allsidig verktøy for ROS-avbildning som kan brukes effektivt in vivo.
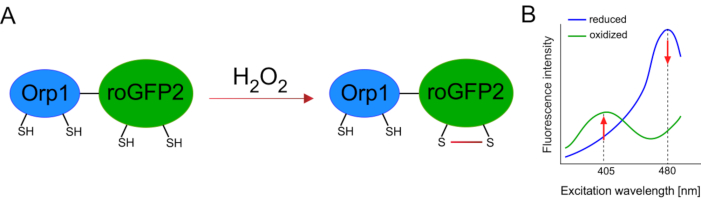
Figur 1: Skjematisk representasjon og eksitasjonsspektra av roGFP2-Orp1. (A) Oksidantoverføring skjer mellom Orp1 og roGFP2 som svar på H2O2, noe som fører til konformasjonsendringer i roGFP2. (B) Eksitasjonsspektraet til roGFP2-Orp1 viser to eksitasjonstopper på 405 nm og 480 nm og enkeltutslippstopp på 515 nm. Ved oksidasjon med H2O2øker eksitasjonen på 405 nm mens eksitasjonen på 480 nm minker. Dette resulterer i en ratiometric avlesning for H2O2 tilstedeværelse. Figuren er modifisert fra Bilan og Belousov (2017)16 og Morgan et al. (2011)25. Klikk her for å se en større versjon av denne figuren.
Danio rerio (sebrafish) modellsystemet har flere fordeler for å bruke genetisk kodede biosensorer. Den optiske gjennomsiktigheten til embryoer og larver muliggjør ikke-invasiv in vivo-avbildning. Nye bildeverktøy utvikles for å oppnå høyere oppløsning og dypere penetrasjon17. Videre er det etablerte verktøy for genetisk manipulasjon (ektopisk mRNA-uttrykk, Tol2 transgenese, etc.) og genomredigering (TALENer, CRISPR / Cas9, etc.), som fremmer generering av transgene dyr18. Etter hvert som sebrafiskembryoene utvikler seg utenfor moren, gir dette systemet ytterligere enklere tilgang og manipulering av embryoene. For eksempel kan encellede injeksjoner og narkotikabehandlinger enkelt gjøres.
Her brukte vi sebrafisk for å midlertidig uttrykke H2O2-spesifikk biosensor roGFP2-Orp1 ved å injisere in vitro-transkribert mRNA. Disse embryoene kan brukes til både in vitro-avbildning av dyrkede nevroner og in vivo-avbildning (figur 2). Vi beskriver en protokoll for dissekering og plating retinal ganglion celler (RGCs) fra sebrafisk embryoer etterfulgt av å vurdereH2O2-nivåer i dyrkede nevroner. Deretter presenterer vi en metode for in vivo-avbildning av roGFP2-Orp1-uttrykkende embryoer og larver ved hjelp av konfokal mikroskopi. Denne tilnærmingen tillater ikke bare å bestemme fysiologiske H2O2-nivåer,men også potensielle endringer som skjer i forskjellige utviklingsstadier eller forhold. Totalt sett gir dette systemet en pålitelig metode for å oppdage H2O2 i levende celler og dyr for å studere rollen som H2O2 i utvikling, helse og sykdom.
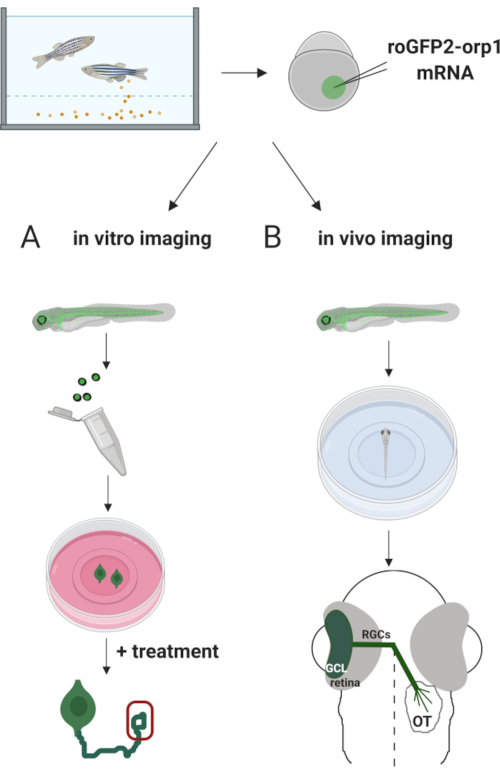
Figur 2. Omriss av den eksperimentelle tilnærmingen. Kort sagt, etter embryosamling injiseres roGFP2-Orp1 mRNA i eggeplommen av encellede sebrafiskembryoer. Utvikling av embryoer kan brukes til både (A) in vitro og (B) in vivo avbildning. (A) GFP-positive embryoer brukes til å dissekere netthinner for RGC-oppsamling ved 34 hkf. Dissosierte RGCer er belagt på PDL/lamininbelagte deksler i ZFCM (+)-medier. Vekstkjegleavbildning kan utføres ettersom RGCer utvider axonene sine etter 6-24 timers plating. Celler kan utsettes for ulike behandlinger for å måle de potensielle endringene i H2O2-nivåer. Her målte viH2O2-nivåer i vekstkjeglene til RGCer (rød). (B) GFP positive embryoer brukes til in vivo-avbildning. I ønsket alder kan embryoer bedøves og monteres på 35 mm glassbunnsretter for konfikal avbildning. Her er embryoer montert ventrally for retinal avbildning. Skjematisk viser retinal utvikling i sebrafisk. RGCer danner ganglion cellelag (GCL), som er det innerste laget i netthinnen. RGC-axoner utvikler seg til optisk nerve for å krysse midtlinjen, og danner optisk chiasm. Deretter vokser RGC-axoner dorsalt for å lage synapser i det optiske tectum i midbrain. Klikk her for å se en større versjon av denne figuren.
