Reaktiva syrearter (ROS) signalering reglerar utvecklingen och funktionen hos nervsystemet1. En viktig cellulär ROS källa är NADPH oxidaser (NOX), som är transmembranproteiner som genererar superoxid och väteperoxid (H2O2)2. NOX enzymer finns i hela centrala nervsystemet (CNS), och NOX-härledda ROS bidrar till neuronal utveckling3,4,5,6. Underhåll och differentiering av neurala stamceller, upprättande av neuronal polaritet, neurit utväxt och synaptisk plasticitet har visat sig kräva tillräckliga nivåer av ROS7,8,9,10,11. Å andra sidan bidrar okontrollerad produktion av ROS av NOXes till neurodegenerativa störningar inklusive Alzheimers sjukdom, multipel skleros och traumatisk hjärnskada12,13,14. Därför är produktion av fysiologiskt relevant ROS avgörande för att upprätthålla hälsosamma förhållanden.
Utveckling av genetiskt kodade biosensorer underlättade upptäckten av cellulära ROS kraftigt. En viktig fördel med genetiskt kodade biosensorer är den ökade temporala och rumsliga upplösningen av ROS-signalen, eftersom dessa sensorer specifikt kan riktas till olika platser. Redoxkänslig GFP (roGFP) är en typ av sådana ROS-biosensorer. RoGFP2-Orp1-varianten upptäcker specifikt H2O2 genom sin Orp1-domän, som är ett glutationperoxiredoxinfamiljprotein från jäst15,16. Oxidationen av Orp1-proteinet överförs till roGFP2 för att ändra dess konformation (figur 1A). Sonden uppvisar två excitationstoppar nära 405 nm och 480 nm, och en enda utsläppstopp vid 515 nm. Vid oxidation förändras fluorescensintensiteten runt excitationstopparna: medan 405 nm excitation ökar minskar 480 nm excitation. Således är roGFP2-Orp1 en ratiometric biosensor, och H2O2-nivåerdetekteras av förhållandet mellan fluorescensintensiteter upphetsade vid två olika våglängder (Figur 1B). Sammantaget är roGFP2-Orp1 ett mångsidigt verktyg för ROS-avbildning som kan användas effektivt in vivo.
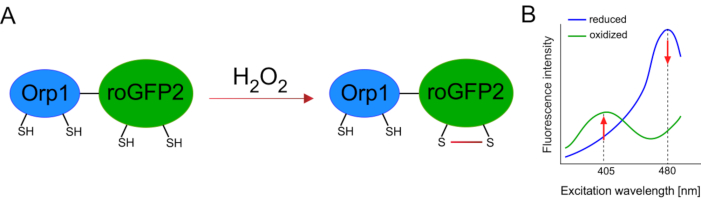
Figur 1: Schematisk representation ochexcitationsspektra av roGFP2-Orp1. (A) Oxidantöverföring sker mellan Orp1 och roGFP2 som svar på H2O2,vilket leder till konformationsförändringar i roGFP2. B)RoGFP2-Orp1:s excitationsspektra uppvisar två excitationstoppar vid 405 nm och 480 nm och en enda utsläppstopp vid 515 nm. Vid oxidation med H2O2ökar excitationen på 405 nm medan 480 nm excitation minskar. Detta resulterar i en ratiometric avläsning för H2O2 närvaro. Siffran har ändrats från Bilan och Belousov (2017)16 och Morgan et al. (2011)25. Klicka här om du vill visa en större version av den här figuren.
Danio rerio (zebrafisk) modellsystem har flera fördelar med att applicera genetiskt kodade biosensorer. Den optiska transparensen hos embryon och larver möjliggör icke-invasiv in vivo-avbildning. Nya bildverktyg utvecklas för att uppnå högre upplösning och djupare penetration17. Dessutom finns det etablerade verktyg för genetisk manipulering (ectopic mRNA-uttryck, Tol2-transgenes, etc.) och genomredigering (TALENs, CRISPR/ Cas9, etc.), vilket främjar genereringen av transgena djur18. När zebrafiskembryon utvecklas utanför modern möjliggör detta system ytterligare enklare åtkomst och manipulering av embryona. Till exempel kan encelliga injektioner och läkemedelsbehandlingar enkelt göras.
Här använde vi zebrafisk för att tillfälligt uttrycka H2O2-specifikabiosensor roGFP2-Orp1 genom att injicera in vitro-transkriberad mRNA. Dessa embryon kan användas för både in vitro-avbildning av odlade nervceller och in vivo-avbildning (figur 2). Vi beskriver ett protokoll för dissekering och plätering retinal ganglion celler (RGCs) från zebrafish embryon följt av bedömning H2O2-nivåer i odlade nervceller. Sedan presenterar vi en metod för in vivo-avbildning av roGFP2-Orp1-uttryckande embryon och larver med konfokal mikroskopi. Detta tillvägagångssätt tillåter inte bara att bestämma fysiologiska H2O2-nivåerutan också potentiella förändringar som inträffar i olika utvecklingsstadier eller förhållanden. Sammantaget ger detta system en tillförlitlig metod för att upptäcka H2O2 i levande celler och djur för att studera H2O2: s roll i utveckling, hälsa och sjukdom.
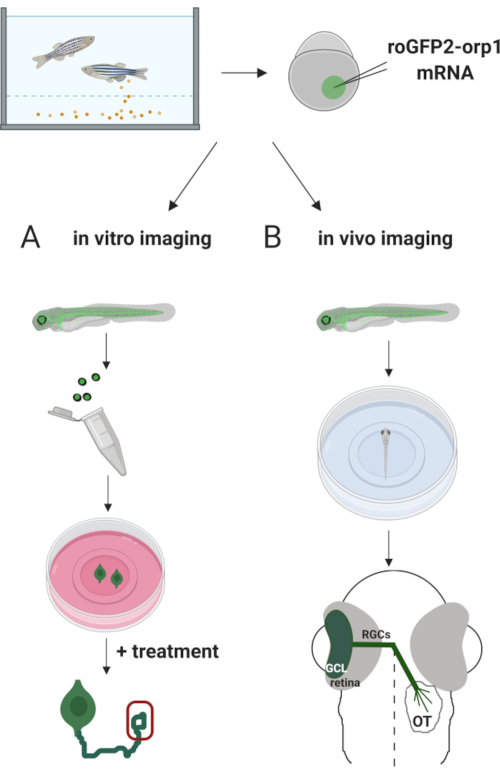
Figur 2. Översikt över det experimentella tillvägagångssättet. Kort, efter embryosamling, injiceras roGFP2-Orp1 mRNA i äggulan av encelliga zebrafiskembryon. Utveckling av embryon kan användas för både (A) in vitro och (B) in vivo imaging. A)GFP-positiva embryon används för att dissekera näthinne för RGC-insamling vid 34 hpf. Dissocierade RGCs pläteras på PDL/lamininbelagda täcken i ZFCM (+) media. Tillväxt kon imaging kan utföras som RGCs utöka sina axons efter 6-24 h plätering. Celler kan utsättas för olika behandlingar för att mäta de potentiella förändringarna i H2O2-nivåer. Här mätte vi H2O2-nivåeri tillväxtkonerna hos RGCs (röd). B)GFP-positiva embryon används för in vivo-avbildning. Vid önskad ålder kan embryon bedövas och monteras på 35 mm glasbottenfat för konfokal avbildning. Här monteras embryon ventrally för retinal avbildning. Schematiska visar retinal utveckling i zebrafisk. RGCs bildar ganglion cell lager (GCL), som är det innersta skiktet i näthinnan. RGC axoner utvecklas till optisk nerv för att korsa mittlinjen, bilda optik chiasm. Sedan växer RGC-axoner dorsally för att göra synapser i det optiska tectumet i midbrain. Klicka här om du vill visa en större version av den här figuren.
