Методика профилирования рибосом (RIBO-seq) была разработана в лаборатории Джонатана Вайсмана в Калифорнийском университете в Сан-Франциско1. По сравнению с другими методами, используемыми для изучения экспрессии генов на трансляционном уровне, RIBO-seq фокусируется на связывании каждой рибосомы с мРНК и предоставляет информацию о ее местоположении и относительном количестве рибосом на транскрипте. Он позволяет контролировать процесс синтеза белка in vivo и может обеспечить одно разрешение и точность кодона, что позволяет измерять плотность рибосом как на отдельной мРНК, так и на всем транскриптоме в клетке. В основе методики RIBO-seq лежит тот факт, что во время трансляции рибосома связывает молекулу мРНК и тем самым защищает погребенный фрагмент транскрипта от переваривания рибонуклеазы. При добавлении рибонуклеазы незащищенная мРНК переваривается, и фрагменты, заключенные рибосомами – обычно длиной ~ 28-30 нт – остаются нетронутыми. Эти фрагменты, называемые рибосомными следами (RF), затем могут быть выделены, секвенированы и нанесены на транскрипт, из которого они произошли, что приводит к обнаружению точного положения рибосом. Фактически, способность рибосомы защищать фрагменты мРНК используется с 1960-х годов для изучения рибосомных сайтов связывания и инициации трансляции (TIS)2,3,4. Однако с развитием технологии глубокого секвенирования RIBO-seq стал золотым стандартом для мониторинга трансляции5, который благодаря взаимодействию с рибосомами может предоставить общегеномную информацию о синтезе белка6. Профилирование рибосом заполнило технологический пробел, который существовал между количественной оценкой транскриптома и протеома6.
Для проведения рибосомного профилирования необходимо получить лизат клеток организма, выросших в исследуемых условиях. Нарушение этих условий во время сбора и лизиса клеток может обеспечить ненадежные данные. Чтобы предотвратить это, обычно используются ингибиторы трансляции, быстрый сбор и мгновенное замораживание в жидком азоте. Клетки могут быть лизированы криогенным измельчением в механическом гомогенизаторе, таком как смеситель мельницы7,8 или бисерный взбиватель9,и путем тритурации через пипетку10 или с помощью иглы11. Буфер лизиса может быть добавлен непосредственно перед или вскоре после измельчения клеток. В нашем протоколе мы используем жидкий азот для предварительного охлаждения раствора и пестиков, а также оксид алюминия в качестве более мягкого подхода к разрушению бактериальной клеточной стенки, что предотвращает сдвиг РНК, часто встречающийся при применении таких методов, как сонификация. После измельчения мы добавляем ледяной буфер лизиса в охлажденный содержимое раствора. Выбор подходящего буфера лизиса важен для получения наилучшего разрешения рибосомных следов. Поскольку ионная сила влияет как на размер ВЧ, так и на точность кадра считывания, в настоящее время рекомендуется использовать лизисные буферы с низкой ионной силой и буферной емкостью, даже если окажется, что буферный состав не влияет на рибосомную занятость на мРНК11,12. Важными компонентами лизисного буфера являются ионы магния, наличие которых предотвращает диссоциацию рибосомных субъединиц и подавляет спонтанные конформационные изменения в бактериальных рибосомах11,13. Ионы кальция также играют значительную роль и необходимы для активности микрококковой нуклеазы (МНазы), используемой в методе профилирования бактериальных рибосом14. Добавление гуанозина 5′-[β,γ-имидо]трифосфата (GMP-PNP), негидролизуемого аналога ГТФ, вместе с хлорамфениколом ингибирует трансляцию во время лизиса15.
При получении лизата его осветляют центрифугированием и делят на две части, каждая для RIBO-seq и высокопроизводительного полного секвенирования мРНК (RNA-seq), поскольку они выполняются одновременно(фиг.1). RNA-seq обеспечивает точку отсчета, которая позволяет сравнивать данные как RIBO-seq, так и RNA-seq во время анализа данных. Исследуемый транслейлом определяется нормализацией рибосомных следов до обилия мРНК16. Данные из RNA-seq также могут помочь идентифицировать артефакты клонирования или секвенирования17.
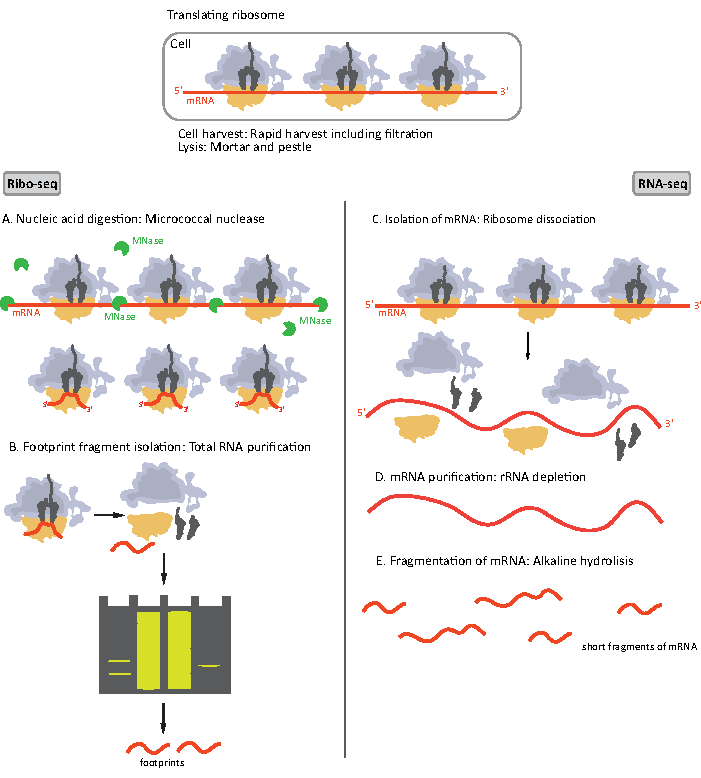
Рисунок 1. Схемы пробоподготовки мРНК для RIBO-seq и RNA-seq. Для подготовки библиотеки RIBO-seq РНК переваривается МНазой (A), с последующим выбором размера РЧ ~28-30 nt длины (B); для РНК-seq РНК выделяют (С), истощают рРНК (D), а полученную мРНК случайным образом фрагментируют на фрагменты различной длины (Е). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Начальные этапы процедуры пробоподготовки для RIBO-seq и RNA-seq отличаются незначительно(рисунок 1). Для рибосомного профилирования лизат должен быть переварен специфической эндонуклеазой, чтобы раздуть молекулы мРНК, не защищенные рибосомами. В стандартных протоколах полученные моносомы восстанавливают ультрацентрифугированием сахарозной подушки или ультрацентрифугированием градиента сахарозы8,14. В этой статье мы показываем, что этот шаг не является необходимым для выделения РЧ, необходимого для RIBO-seq у бактерий, аналогично для эукариотических клеток18,и что выбор размера фрагментов мРНК соответствующей длины из полиакриламидного геля является достаточным.
Для РНК-seq мРНК получается путем истощения рРНК из общей РНК – молекулы рРНК гибридизируются с биотинилированными олигонуклеотидными зондами, которые связываются с магнитными шариками, покрытыми стрептавидином. Комплексы рРНК-олигонуклеотид-шарики затем удаляются из образца магнитом, в результате чего рРНК истощается образцом19,20. Очищенные молекулы мРНК затем случайным образом фрагментируются щелочным гидролизом. Полученные фрагменты мРНК, а также рибосомные следы преобразуются в библиотеки кДНК и подготавливаются к глубокому секвенированию(рисунок 2). Это включает в себя восстановление концов, необходимое после щелочного гидролиза (для мРНК) и ферментативного пищеварения (для RF): дефосфорилирование 3′ концов с последующим фосфорилированием 5′ концов. Следующими шагами являются лигирование адаптеров и обратная транскрипция для создания вставок кДНК, обрамленных последовательностями, необходимыми для секвенирования следующего поколения (NGS) с использованием платформы Illumina. Последняя фаза подготовки библиотеки представляет собой реакцию ПЦР, в которой конструкции усиливаются и маркируются штрих-кодами для конкретных образцов, чтобы обеспечить мультиплексирование и секвенирование различных образцов на одном канале. Перед секвенированием качество и количество библиотек оцениваются с помощью высокочувствительного электрофореза ДНК на кристалле. Затем библиотеки cDNA с соответствующими параметрами могут быть объединены в пул и секвенированы. Секвенирование может выполняться на различных платформах Illumina, таких как MiSeq, NextSeq или HighSeq, в зависимости от количества библиотек, требуемой длины чтения и глубины секвенирования. После секвенирования проводится биоинформатический анализ.

Рисунок 2. Подготовка библиотеки. Подготовка библиотеки включает в себя ремонт концов, перевязку адаптеров, обратную транскрипцию и усиление штрих-кодированием. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Профилирование рибосом является универсальным методом, который может быть легко модифицирован и скорректирован в соответствии с научным вопросом. Первоначально он использовался в дрожжах1,но вскоре после этого он был применен к бактериальным клеткам21, а также к эукариотическим модельным организмам, включая мышь10,рыбку данио22,плодовуюмуху 23 и Arabidopsis thaliana24. Он также использовался для изучения различных типов рибосом: цитоплазматических, митохондриальных25,26 и хлоропластов27,28. У эукариот RIBO-seq обычно адаптируется и уточняется для исследования конкретных аспектов трансляции, включая инициацию10,11,29,30,31,32,удлинение1,10,11,31,33,рибосомное сваливание33 и изменение конформации33. Большинство модификаций предполагают использование различных ингибиторов трансляции. Однако на бактериях аналогичные исследования было трудно провести из-за нехватки ингибиторов с требуемым механизмом действия34. Наиболее часто используемым ингибитором трансляции у бактерий является хлорамфеникол (CAM), который связывается с центром пептидилтрансферазы (PTC) и предотвращает правильное позиционирование аминоацил-тРНК в А-сайте. В результате CAM предотвращает образование пептидной связи, что приводит к остановке удлиняющихся рибосом35. Другими примерами ингибиторов трансляции у бактерий являются тетрациклин (TET)36,ретапамулин (RET)34 и Onc11237, которые использовались для исследования сайтов инициации трансляции. ТЕТ, который предотвращает доставку тРНК в рибосому путем прямого перекрытия с антикодоном стволовым контуром тРНК в А-месте, первоначально применялся для проверки результатов, полученных от лечения CAM, поскольку они оба являются антибиотиками, ингибируя удлинение трансляции38. Было обнаружено, что TET обнаруживает первичный TIS, однако не смог выявить внутренний TIS36. RET связывается в PTC бактериальной рибосомы и предотвращает образование первой пептидной связи путем вмешательства в элонгатор аминоацил-тРНК в месте А. Применение RET приводит к остановке рибосом как на первичном, так и на внутреннем TISs34. Onc112, богатый пролином антимикробный пептид, связывается в выходном туннеле и блокирует связывание аминоацил-тРНК в рибосомном участке А. В результате Onc112 предотвращает вход комплексов инициации в фазуудлинения 37.
Основной информацией, которую обеспечивает профилирование рибосом, является плотность рибосом и их положение на мРНК. Он был успешно применен для исследования дифференциальной экспрессии генов на уровне трансляции в различных условиях роста1,6,измерения поступающей эффективности1,38,39 и обнаружения событий регуляции трансляции, таких как рибосомная пауза10. RIBO-seq также позволяет раскрыть трансляцию аннотированных ncRNA, псевдогенов и неаннотированных малых открытых кадров чтения (ORF), что приводит к идентификации новых и/или очень коротких генов, кодирующих белки10,12,22,30,37. В таких случаях RIBO-seq может точно настроить и улучшить аннотацию генома. Обладая высокой чувствительностью к идентификации переведенных ОРФ и его количественной природой, профилирование рибосом может также служить прокси для определения протеома или в оказании помощи протеомным исследованиям31,34,39. Путем картирования TIS, профилирование рибосом выявляет N-терминально расширенные и усеченные изоформы известных белков10,32. RIBO-seq также был адаптирован для изучения ко-трансляционного сворачивания белков14,21,24. Этот метод позволяет измерять скорости удлинения1,10,39 или скорость декодирования отдельных кодонов6 и помогает в разработке количественных моделей трансляции17. Метод профилирования рибосом также способен обеспечить механистическое понимание паузы рибосомы у бактерий7,15,17,сдвига кадра40,стоп-кодона через21,дефектов терминации/рециркуляции41,42 и рибосомных конформационных изменений33 у эукариот. RIBO-seq также был адаптирован для изучения влияния специфических трансдействующих факторов на трансляцию, таких как миРНК6 и РНК-связывающие белки у эукариот16,43. Однако важно признать, что экспериментальная конструкция и полученная разрешение RIBO-seq определяют объем информации, которая может быть извлечена из полученных данных секвенирования12.
