Epilepsi, den fjerde vanligste familien av nevrologiske lidelser i USA1, er preget av en ubalanse av eksitatorisk og hemmende drivkraft i CNS som fører til tilbakevendende anfall. Feberbeslag (FS) eller feberrelaterte anfall kan forekomme i befolkningen generelt, oftest hos barn så tidlig som 3 måneder opp til 6-7 år. Men hos enkelte individer med genetiske mutasjoner, oftest i et natriumkanalgen, kan FS vedvare utover 7 år i voksen alder. Denne tilstanden kalles feberanfall pluss eller FS+. Raske fremskritt innen genomsekvensering har identifisert over 1300 mutasjoner i det humane natriumionkanalgenet SCN1A, noe som gjør det til et hotspot for epilepsimutasjoner. SCN1A-mutasjoner har vært knyttet til et bredt spekter av anfallsforstyrrelser, inkludert feberbeslag (FS), genetisk epilepsi med feberanfall pluss (GEFS+) og Dravet syndrom (DS)2,3,4,5,6. Omtrent 20% av SCN1A missense mutasjoner fører til GEFS + 5,7,8. Pediatrisk historie med kompleks eller langvarig FS i barndommen kan senere utvikle seg til mer svekkende former for epilepsi som temporal lobe epilepsi (TLE)9,10,11. Dravet syndrom oppstår på grunn av avkortingsmutasjoner eller tap av funksjonsmutasjoner i SCN1A og er en alvorlig form for intractable epilepsi, med barndoms utbrudd av feberanfall som utvikler seg til ildfaste anfall, og er ofte forbundet med kognitive, utviklingsmessige og motoriske svekkelser2,5,12 . Siden mange individer med GEFS+ og/eller DS viser feberanfall, blir det viktig å utvikle nye terapier for bedre å bekjempe disse anfallsforstyrrelsene.
Dyremodeller av SCN1A assosiert epilepsi har vist seg uvurderlig i karakterisering av ulike typer anfall (febril vs generalisert) og dissekering av nevronmekanismen for anfallsgenerasjon13,14,15,16,17,18. Mens studien av spontane anfall via EEG / EMG-opptak i gnagerhjerner er godt etablert og er et veldig nyttig verktøy, har bare noen få studier forsøkt å etterligne feberbeslag i musemodeller14,16,19,20,21,22,23 . Tidligere studier har brukt en stråle med oppvarmet tørr luft, eller en metakrylatsylinder utstyrt med et termisk system, eller varmelamper med temperaturregulator i lukkede testarenaer9,16,21,22,23,24 for å indusere anfall via hypertermi. For å øke kroppstemperaturen i et mer kontrollert miljø, bruker protokollen som er beskrevet her et spesialbygget kammer med et temperaturkontrollert varmesystem som tillot reproduserbare økningshastigheter i kroppstemperaturen til en mus inne i kammeret. Varmekammeret ble konstruert av tre (lengde 40 cm x bredde 34 cm x høyde 31 cm) og ble utstyrt med en digital temperaturregulator med K termokobling. En liten aksialvifte utstyrt med en varmeapparat på baksiden av kammeret leder oppvarmet luft inn i kammeret regulert av en digital temperaturregulator. Dette tvungne luftvarmesystemet gjør det mulig å kontrollere hastigheten kammertemperaturen øker med. (Figur 1A,B). K-termoelementet inne i trevarmekammeret sender tilbakemelding til den digitale temperaturregulatoren for å opprettholde konstante temperaturer inne i esken under analysen. Hvis du stiller inn temperaturen på den digitale temperaturregulatoren, kan den elektriske viften sende oppvarmet tvungen luft gjennom ventiler for å varme opp kammeret jevnt (figur 1A). Frontpanelet på varmekammeret er et klart plexiglassark for å muliggjøre enkel videoopptak av forsøkene.
Voksne (P30-P40) mus, heterozygote for en missense mutasjon i SCN1A som får GEFS+ og et likt antall villtype kullkamerater til å fungere som kontrollgruppen, ble valgt for hvert eksperiment. Dyr, både menn og kvinner, som ble brukt i disse studiene veide minst 15 g, da villmus som veide mindre var mer følsomme for varmeinduserte anfall enn tyngre dyr i samme alder. I pilotstudien ble både mutant- og villtypemus observert for å oppsøke de kjøligere hjørnene av kammeret på baksiden og forble der i lengre perioder. For å omgå dette ble effektiv gulvstørrelse inne i varmekammerets testarena redusert til lengde 16,5 cm x bredde 21,5 cm x høyde 27,5 cm ved å plassere en treblokk B (mål 20 cm x 8 cm x 7,2 cm) på høyre side av kammeret (figur 1A). Varmekammeret ble konstruert av 1,9 cm tykt kryssfiner (lengde 40 cm x bredde 34 cm x høyde 31 cm) dekket med hvitt laminat og utstyrt med en digital temperaturregulator med K termokobling. Kammerveggenes laminatoverflate er ugjennomtrengelig og kan enkelt desinfiseres mellom forsøk ved å tørke av med 70% etanol. Temperaturen på varmekammeret ble i utgangspunktet satt til 50 °C og forvarmet i minst 1 time før starten av eksperimentet, for å sikre jevn oppvarming inne i kammeret. Hver mus var utstyrt med et rektal termometer for kontinuerlig overvåking av kroppstemperatur gjennom hele eksperimentet. En enkelt mus ble plassert i kammeret om gangen og temperaturen ble holdt ved 50 °C mellom 1.–10. Temperaturen ble deretter hevet til 55 °C i 11.-20. minutt, og til slutt hevet til 60 °C i 21.-30. Dette resulterte i en reproduserbar økning i musens kroppstemperatur (figur 2A). Hver prøveversjon ble filmet, og atferdsanalyse ble utført offline.
Varmeprotokollen kan enkelt modifiseres for å endre den opprinnelige temperaturen på varmekammeret og hastigheten som kammeret oppvarmes, noe som igjen endrer hvor raskt kroppstemperaturen til musen er forhøyet under analysen. Dermed gir denne metoden mer fleksibilitet i forhold til tradisjonelle metoder for å sette opp atferdsskjermer som involverer varmeinduserte anfall. Den varmeinduserte anfallsprotokollen kan også brukes til å screene for antiepileptika som gjør mutantmus mer motstandsdyktige mot varmeinduserte anfall eller øker terskeltemperaturen der anfall observeres. På samme måte kan gunstige effekter av restriktive diettregimer som keto-diett på varmeinduserte anfall undersøkes i normale chow-matet vs keto-matede mus.
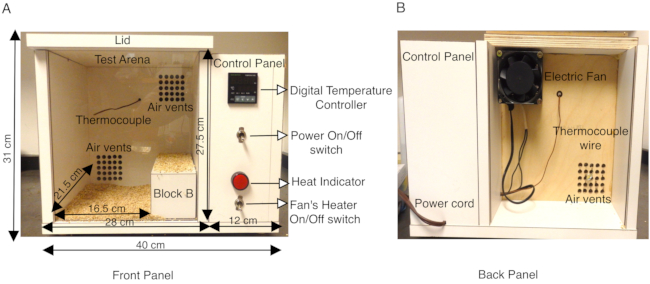
Figur 1: Beskrivelse av det spesialbygde musevarmekammeret. (A) Frontpanelet på tremusens varmekammer viser sidekontrollpanelet som inneholder Power ON/OFF-bryteren som slår på digital temperaturregulator, K-termokobling, vifteovnens PÅ/AV-bryter og varmeindikator. De ytre dimensjonene på boksen og den indre testarenaen vises i cm. En treblokk B som brukes til effektivt å redusere testarenaoverflaten, vises også. Bunnen av testarenaen er dekket med cob sengetøy for å forhindre at mus kommer direkte i kontakt med oppvarmede treflater. (B) Bakpanelet på varmekammeret viser viften montert på toppluftventilen og strømledningen for å levere strøm til kammeret. Dette tallet er modifisert fra figur 3 i Das et al., 2021, eNeuro14. Klikk her for å se en større versjon av denne figuren.
