Ved å levere og fjerne proteiner til og fra plasmamembranen kontrollerer membranproteinhandel i nevroner plasmamembranutstyret med reseptorer samt ionkanaler og regulerer derfor nevronfunksjonen. Feilregulering eller defekter i proteinhandel har vanligvis skadelige effekter på celler og resulterer i nevronal degenerasjon. Hos mennesker kan dette forårsake nevrodegenerative sykdommer som Alzheimers og Parkinsons sykdom eller Retinitis pigmentosa1. Fotoreseptorer i det sammensatte øyet til Drosophila melanogaster har blitt et in vivo-modellsystem for å studere membranproteinhandel2. Dette skyldes ikke bare den genetiske allsidigheten til Drosophila som tillater effektive genetiske skjermer, men også fordi alle viktige komponenter i den lysabsorberende fotoreseptormembranen er preget i detalj og effektive mikroskopiske teknikker er tilgjengelige som kan brukes på flueøyet. Disse teknikkene er fokus i denne artikkelen.
I Drosophila fotoreseptorceller danner den apikale plasmamembranen en tettpakket stabel med mikrovilli langs den ene siden av cellen, kalt rhabdomere. Rhabdomeres av fotoreseptorceller R1-6 er ordnet i et karakteristisk trapesformet mønster mens fotoreseptorceller R7 og R8 danner en enkelt rhabdomere i midten av denne trapesformet3. Membranproteinhandel er nødvendig for en regulert omsetning av rhabdomerale membranproteiner som rhodopsin og den lysaktiverte TRP (forbigående reseptorpotensial) og TRPL (TRP-lignende) ionkanaler for å sikre riktig mengde av disse fototransduksjonsproteinene i rhabdomere. Fotoreseptormembranproteiner syntetiseres i det endoplasmatiske retikulumet og transporteres via Golgi-apparatet til rhabdomere. Etter aktivering av rhodopsin ved lys, kan et rhodopsinmolekyl enten inaktiveres ved absorpsjon av en annen foton eller kan fjernes fra rhabdomere ved klarrinmediert endokytose. Endocytosed rhodopsin blir enten degradert i lysosomet eller resirkuleres tilbake til rhabdomere 4,5. Ionkanalen TRPL er også internalisert etter aktivering av fototransduksjonskaskaden og gjennomgår en lysavhengig translokasjon mellom rhabdomere (hvor den ligger når fluer holdes i mørket) og et ER-beriket oppbevaringsrom i cellekroppen (som den transporteres innen flere timer etter belysning) 6,7,8,9,10 . I motsetning til endokytosed rhodopsin, blir bare små mengder TRPL degradert via den endolysosomale banen, og flertallet lagres intracellulært i stedet og resirkuleres tilbake til rhabdomere ved mørk tilpasning6. TRPL kan dermed brukes til å analysere lysutløst smugling av plasmamembranproteiner. Drosophila fotoreseptorceller brukes også til å studere nevronal degenerasjon. Photoreceptor celle degenerasjon bestemmes ofte ved å vurdere strukturen av rabdomerer, som oppløses som følge av degenerative prosesser5.
For å studere subcellulær lokalisering av TRPL og rhodopsin i fotoreseptorceller eller fotoreseptorcelledegenerering, er det brukt to fluorescensmikroskopimetoder som varierer med hensyn til analysehastighet og oppløsning her. En veldig rask, ikke-invasiv metode som kan brukes til genetiske skjermer, men med en begrenset romlig oppløsning er påvisning av fluorescens i det dype pseudopupilet (DPP). DPP er et optisk fenomen av leddyr sammensatte øyne hvis geometriske opprinnelse har blitt forklart i detalj av Franceschini og Kirschfeld i 197111. Kort sagt, på flere optiske plan under netthinnen overlegg-bilder av rhabdomeres fra tilstøtende ommatidia kan observeres. På et fokalplan gjennom midten av øyets krumning danner disse overliggende projeksjonene et bilde som ligner den trapesformede utformingen av rhabdomeres i et enkelt ommatidium bare størrelsesorden større. Dette fenomenet kan også observeres uavhengig av eksogent uttrykk for fluorescensproteiner (f.eks. TRPL::eGFP8), noe som likevel gjør DPP enklere å oppdage (figur 1A-A‘)12. En annen ikke-invasiv metode er vanninnlevelsesmikroskopi som er avhengig av avbildning av fluorescerende merkede proteiner etter optisk nøytralisering av øynenes dioptriske apparat med vann (figur 1B-C‘)12. Ved hjelp av vann nedsenkningsmetoden kan den relative mengden TRPL::eGFP i rhabdomeres eller cellekropp vurderes kvantitativt for individuelle fotoreseptorceller. Videre kan ikke-translokerende fluorescensmerkede proteiner brukes til å evaluere rabdomeral integritet og for å bestemme tidsforløpet for en potensiell degenerasjon på en kvantitativ måte, som beskrevet her.
Mens opptak av DPP er den desidert enkleste og raskeste av disse metodene å utføre, er den romlige oppløsningen av data de genererer begrenset. I tillegg er det mange grunner til at en DPP kan være fraværende, som ikke nødvendigvis er merkbar av DPP-bildebehandling selv. Siden DPP representerer en oppsummering av flere ommatidia, går informasjon om individuelle celler tapt. Dermed tjener lavoppløselig DPP-avbildning en viktig funksjon i screening av et stort antall fluer, men bør generelt følges av opptak med høyere oppløsning ved hjelp av vanninnlevelsesmikroskopi. Vann nedsenkningsmikrografer tillater tolkninger om individuelle celler, utviklingsfeil, øyemorfologi, proteinfeillokalisering eller retinal degenerasjon samt kvantifisering av disse effektene. Denne protokollen beskriver disse to teknikkene i detalj.
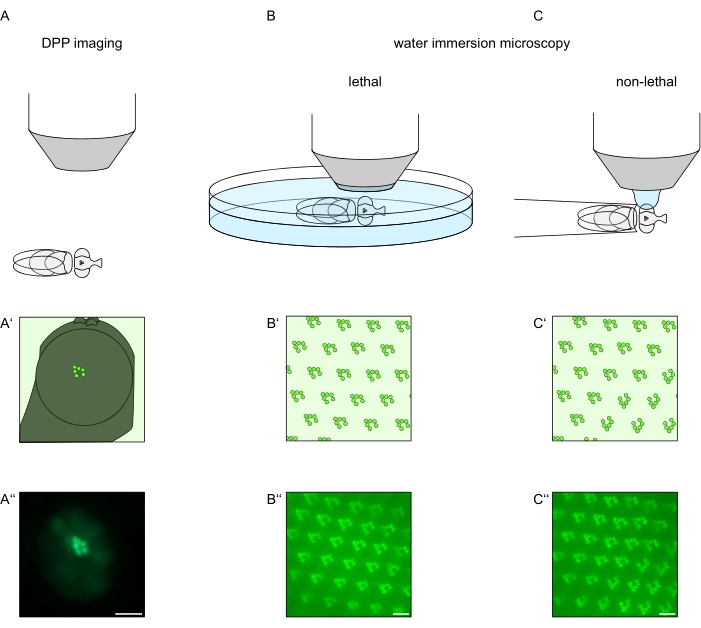
Figur 1: Oversikt over mikroskopivariasjoner for Drosophila-øyet presentert i denne protokollen. Skjematiske representasjoner og eksemplariske mikrografer av (A-A”) fluorescerende dyp pseudopupil (DPP) avbildning, (B-B”) dødelig vanninnlevelsesmikroskopi av fluorescerende rabdomerer, og (C-C”) ikke-dødelige vanndråpemikroskopi av fluorescerende rhabdomere. Skalastang (A”): 100 μm. Skalastolper (B”–C”): 10 μm. Tallet er endret fra referanse13. Klikk her for å se en større versjon av denne figuren.
