En délivrant et en éliminant les protéines vers et depuis la membrane plasmique, le trafic de protéines membranaires dans les neurones contrôle l’équipement de la membrane plasmique avec des récepteurs ainsi que des canaux ioniques et, par conséquent, régule la fonction neuronale. Une mauvaise régulation ou des défauts dans le trafic des protéines ont généralement des effets néfastes sur les cellules et entraînent une dégénérescence neuronale. Chez l’homme, cela peut provoquer des maladies neurodégénératives telles que la maladie d’Alzheimer et la maladie de Parkinson ou la rétinite pigmentaire1. Les photorécepteurs dans l’œil composé de Drosophila melanogaster sont devenus un système modèle in vivo pour l’étude du trafic de protéines membranaires2. Cela n’est pas seulement dû à la polyvalence génétique de la drosophile qui permet des criblages génétiques efficaces, mais aussi parce que tous les composants essentiels de la membrane photoréceptrice absorbant la lumière sont caractérisés en détail et que des techniques microscopiques efficaces sont disponibles qui peuvent être appliquées à l’œil de la mouche. Ces techniques sont au centre de cet article.
Dans les cellules photoréceptrices de la drosophile, la membrane plasmique apicale forme une pile dense de microvillosités le long d’un côté de la cellule, appelée rhabdomere. Les rhabdomeres des cellules photoréceptrices R1-6 sont disposées selon un motif trapézoïdal caractéristique tandis que les cellules photoréceptrices R7 et R8 forment un seul rhabdomere au centre de ce trapèze3. Le trafic de protéines membranaires est nécessaire pour un renouvellement régulé des protéines de la membrane rhabdomerale telles que la rhodopsine et les canaux ioniques TRP (potentiel récepteur transitoire) et TRPL (TRP-like) activés par la lumière afin d’assurer la quantité appropriée de ces protéines de phototransduction dans le rhabdomere. Les protéines de la membrane photoréceptrice sont synthétisées dans le réticulum endoplasmique et transportées via l’appareil de Golgi vers le rhabdomere. Après l’activation de la rhodopsine par la lumière, une molécule de rhodopsine peut soit être inactivée par absorption d’un deuxième photon, soit être retirée du rhabdomere par endocytose médiée par la clathrine. La rhodopsine endocytosée se dégrade dans le lysosome ou est recyclée en rhabdomere 4,5. Le canal ionique TRPL est également internalisé après activation de la cascade de phototransduction et subit une translocation dépendante de la lumière entre le rhabdomere (où il est situé lorsque les mouches sont maintenues dans l’obscurité) et un compartiment de stockage enrichi en ER dans le corps cellulaire (vers lequel il est transporté en quelques heures après l’illumination)6,7,8,9,10 . Contrairement à la rhodopsine endocytosée, seules de petites quantités de TRPL sont dégradées par la voie endolysosomale, et la majorité est stockée intracellulairement à la place et recyclée dans le rhabdomere lors de l’adaptation sombre6. TRPL peut ainsi être utilisé pour analyser le trafic déclenché par la lumière des protéines de la membrane plasmique. Les cellules photoréceptrices de la drosophile sont également utilisées pour étudier la dégénérescence neuronale. La dégénérescence des cellules photoréceptrices est souvent déterminée en évaluant la structure des rhabdomeres, qui se désintègrent à la suite de processus dégénératifs5.
Afin d’étudier la localisation subcellulaire de la TRPL et de la rhodopsine dans les cellules photoréceptrices ou la dégénérescence des cellules photoréceptrices, deux méthodes de microscopie à fluorescence qui diffèrent en termes de vitesse d’analyse et de résolution ont été appliquées ici. Une méthode très rapide et non invasive qui peut être utilisée pour les criblages génétiques mais avec une résolution spatiale limitée est la détection de fluorescence dans le pseudopieux profond (DPP). Le DPP est un phénomène optique d’yeux composés d’arthropodes dont l’origine géométrique a été expliquée en détail par Franceschini et Kirschfeld en 197111. En bref, sur plusieurs plans optiques sous la superposition de la rétine, des images de rhabdomeres d’ommatidies adjacentes peuvent être observées. Sur un plan focal passant par le centre de la courbure de l’œil, ces projections superposées forment une image qui ressemble à la disposition trapézoïdale des rhabdomeres dans un seul ommatidium seulement des ordres de grandeur plus grands. Ce phénomène peut également être observé indépendamment de l’expression exogène des protéines de fluorescence (par exemple TRPL::eGFP8), ce qui rend néanmoins le DPP plus facile à détecter (Figure 1A-A”)12. Une deuxième méthode non invasive est la microscopie par immersion dans l’eau qui repose sur l’imagerie de protéines marquées par fluorescence après avoir neutralisé optiquement l’appareil dioptrique des yeux avec de l’eau (Figure 1B-C”)12. En utilisant la méthode d’immersion dans l’eau, la quantité relative de TRPL::eGFP dans les rhabdomeres ou le corps cellulaire peut être évaluée quantitativement pour les cellules photoréceptrices individuelles. En outre, les protéines marquées par fluorescence non translocalisantes peuvent être utilisées pour évaluer l’intégrité rhabdomerale et pour déterminer l’évolution temporelle d’une dégénérescence potentielle de manière quantitative, comme décrit ici.
Bien que les enregistrements du DPP soient de loin les plus faciles et les plus rapides de ces méthodes, la résolution spatiale des données qu’ils génèrent est limitée. En outre, il existe de nombreuses raisons pour lesquelles un DPP peut être absent, qui ne sont pas nécessairement discernables par l’imagerie DPP elle-même. Étant donné que le DPP représente une somme de plusieurs ommatidies, des informations sur les cellules individuelles sont perdues. Ainsi, l’imagerie DPP à basse résolution joue un rôle important dans le dépistage d’un grand nombre de mouches, mais devrait généralement être suivie d’enregistrements à plus haute résolution par microscopie à immersion dans l’eau. Les micrographies à immersion dans l’eau permettent des interprétations sur les cellules individuelles, les défauts de développement, la morphologie des yeux, la mauvaise localisation des protéines ou la dégénérescence rétinienne ainsi que la quantification de ces effets. Le présent Protocole décrit ces deux techniques en détail.
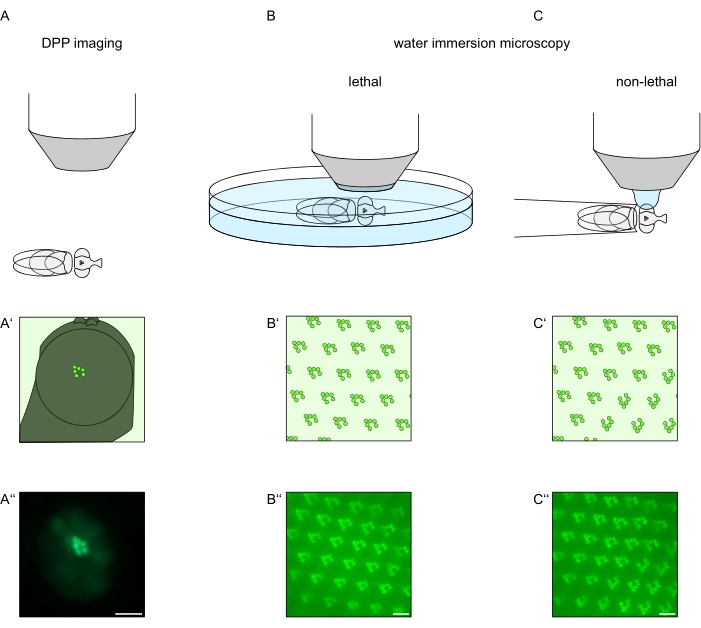
Figure 1 : Aperçu des variations microscopiques de l’œil de drosophile présentées dans ce protocole. Représentations schématiques et micrographies exemplaires de l’imagerie fluorescente par pseudopieux profond (DPP) (A-A”), de la microscopie par immersion dans l’eau létale (B-B”) des rhabdomeres fluorescents et de la microscopie à goutte d’eau non létale (C-C”) des rhabdomeres fluorescents. Barre d’échelle (A”): 100 μm. Barres d’échelle (B”–C”): 10 μm. La figure a été modifiée par rapport à la référence13. Veuillez cliquer ici pour voir une version agrandie de cette figure.
