Une myriade de modèles sont disponibles pour étudier la progression du cancer, chacun d’eux étant unique et représentant un sous-type de cette maladie complexe. Chaque modèle fournit des informations uniques et précieuses sur la biologie du cancer et a amélioré les moyens d’imiter l’état réel de la maladie. Les lignées cellulaires établies cultivées en monocouche ont fourni des informations précieuses sur les processus vitaux in vitro, tels que la prolifération, l’invasivité, la migration et l’apoptose1. Bien que la culture cellulaire bidimensionnelle (2D) ait été l’outil traditionnel pour étudier la réponse des cellules de mammifères à plusieurs perturbations environnementales, l’extrapolation de ces résultats pour prédire les réponses au niveau tissulaire ne semble pas suffisamment convaincante. La principale limitation des cultures 2D est que le microenvironnement créé diffère largement de celui du tissu mammaire lui-même2. La culture 2D manque de l’interaction des cellules avec la matrice extracellulaire, ce qui est vital pour la croissance de tout tissu. De plus, les forces de traction subies par la cellule dans les cultures monocouches entravent la polarité de ces cellules, modifiant ainsi la signalisation et le comportementdes cellules 3,4,5. Les systèmes de culture tridimensionnels (3D) ont ouvert une nouvelle voie dans le domaine de la recherche sur le cancer avec leur capacité à imiter les conditions in vivo in vitro. De nombreux indices microenvironnementaux cruciaux qui sont perdus dans la culture cellulaire 2D pourraient être rétablis en utilisant des cultures 3D de matrice extracellulaire riche en laminine (lrECM)6.
Diverses études ont identifié l’importance du microenvironnement tumoral dans la cancérogenèse 7,8. Les facteurs associés à l’inflammation constituent une partie importante du microenvironnement. Le facteur d’activation plaquettaire (PAF) est un médiateur phospholipide sécrété par diverses cellules immunitaires qui médie les réponses immunitairesmultiples 9,10. Des niveaux élevés de FAP sont sécrétés par différentes lignées cellulaires de cancer du sein et sont associés à une prolifération accrue11. Des études de notre laboratoire ont montré que la présence prolongée de PAF dans les cultures acineuses entraîne la transformation des cellules épithéliales du sein12. PAF active le récepteur PAF (PAFR), activant l’axe de signalisation PI3K/Akt13. Le DPAP est également associé à l’EMT, à l’invasion et aux métastases14.
Le présent protocole démontre un système modèle pour étudier la transformation induite par le PAF, en utilisant des cultures 3D de cellules épithéliales du sein, comme cela a été décrit précédemment par Chakravarty et al.12. Les cellules épithéliales mammaires cultivées sur la matrice extracellulaire (cultures 3D) ont tendance à former des sphéroïdes polarisés arrêtés par croissance. Ceux-ci sont appelés acini et ressemblent étroitement aux acini du tissu mammaire, la plus petite unité fonctionnelle de la glande mammaire, in vivo15. Ces sphéroïdes (Figure 1A,B) sont constitués d’une monocouche de cellules épithéliales polarisées étroitement tassées entourant une lumière creuse et fixées à la membrane basale (Figure 1C). Ce processus de morphogenèse a été bien décrit dans la littérature16. Lorsqu’elles sont ensemencées sur lrECM, les cellules subissent une division et une différenciation pour former un groupe de cellules, qui se polarisent ensuite à partir du jour 4. Au jour 8, les acini sont constitués d’un groupe de cellules polarisées qui sont en contact direct avec la matrice extracellulaire et d’un groupe de cellules non polarisées enfermées dans les cellules polarisées externes, sans contact avec la matrice. Ces cellules non polarisées sont connues pour subir l’apoptose au jour 12 de la culture, formant une lumière creuse. Au jour 16, les structures arrêtées par la croissance sont formées16.
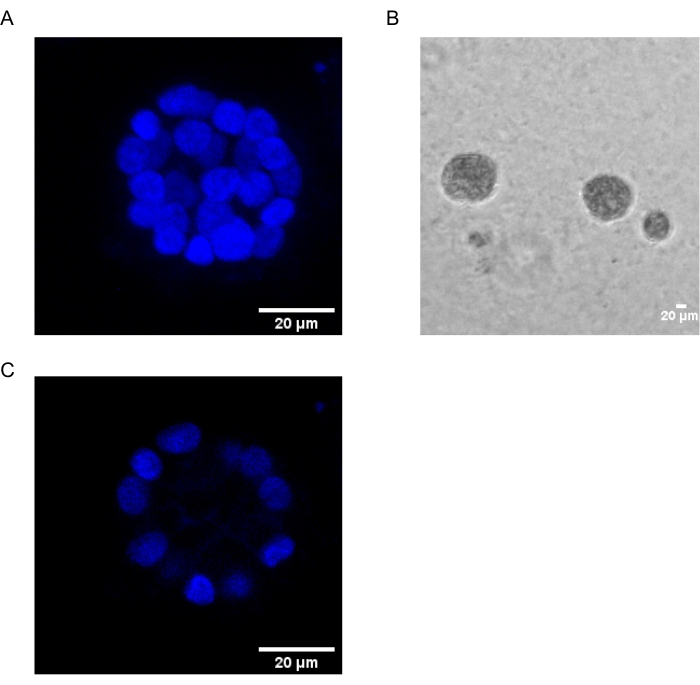
Figure 1 : Noyaux de cellules dans les acini colorés avec une coloration nucléaire. (A) Construction 3D des acini. (B) Image à contraste de phase de MCF10A acini cultivée sur Matrigel pendant 20 jours. (C) La section la plus centrale montre la présence d’une lumière creuse. Barre d’échelle = 20 μm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
Contrairement aux cultures 2D, les cultures acineuses aident à distinguer les cellules normales et transformées par des changements morphologiques apparents. Les cellules épithéliales mammaires non transformées forment des acini avec une lumière creuse, imitant les acini mammaires humains normaux. Ces sphéroïdes, lors de la transformation, présentent une morphologie perturbée caractérisée par une perte importante de polarité (une des caractéristiques du cancer), l’absence de lumière ou une perturbation de la lumière creuse (due à l’évasion de l’apoptose) qui peut être induite en raison de la dérégulation de divers gènes17,18,19,20 . Ces transformations peuvent être étudiées à l’aide de techniques couramment utilisées telles que l’immunofluorescence. Ainsi, le modèle de culture cellulaire 3D peut fonctionner comme une méthode simple pour étudier le processus de morphogenèse acineuse du sein et de cancérogenèse mammaire. L’établissement d’un système de culture 3D pour comprendre l’effet d’un médiateur phospholipidique, PAF, aidera au criblage préclinique de médicaments à haut débit.
Ce travail a adapté le protocole de culture 3D ‘on top’ 16,21 pour étudier la transformation induite par PAF 22. Les changements phénotypiques induits par l’exposition des acini au médiateur phospholipide ont été étudiés par immunofluorescence. Divers marqueurs de polarité et de transition épithéliale à mésenchymateuse (EMT)12,16 ont été utilisés dans l’étude. Le tableau 1 mentionne leur localisation normale et leur phénotype attendu lors de la transformation.
| Anticorps | Marques | Localisation normale | Phénotype transformé |
| α6-Intégrine | Basolatéral | Basal avec faible coloration latérale | Forte tache latérale / apicale |
| β-Caténine | Jonction cellule-cellule | Basolatéral | Localisation anormale / nucléaire ou cytoplasmique |
| Vimentin | Emt | Présence absente / faible | Réglementation à la hausse |
Tableau 1 : Marqueurs utilisés dans l’étude. Différents marqueurs utilisés avec leur localisation en présence et en l’absence de traitement PAF.
Cette méthode peut être mieux utilisée pour étudier / cribler des médicaments plausibles et des gènes cibles pour divers sous-types de cancer du sein. Cela peut fournir des données de réponse aux médicaments plus proches du scénario in vivo , ce qui contribue à un développement de médicaments plus rapide et plus fiable. En outre, ce système peut être utilisé pour étudier la signalisation moléculaire associée à la réponse aux médicaments et à la résistance aux médicaments.
