The cellular activity evaluation of the adjuvants OP-D and NOD was completed in vitro according to the protocol. L929 fibroblasts are a useful screening model for the in vitro toxicity testing of NOD (Figure 1). The quantification of inflammatory cytokine levels in the spleen can help researchers better understand the immune response (Figure 2). Monitoring CTLs with ELISpot is the gold standard for assessing antigen-specific T-cell immunity in clinical trials and for screening vaccine candidates (Figure 3). The increased uptake of an antigen by DCs can elicit enhanced adaptive immune responses (Figure 4). Macrophages play an important role in presenting antigens to T cells, as well as inducing other antigen-presenting cells to express costimulatory molecules, thereby initiating adaptive immune responses (Figure 5). The above experimental results were published by Tong et al., and the different antibody responses in vivo and protection efficiency against mice can be found in the original article14.
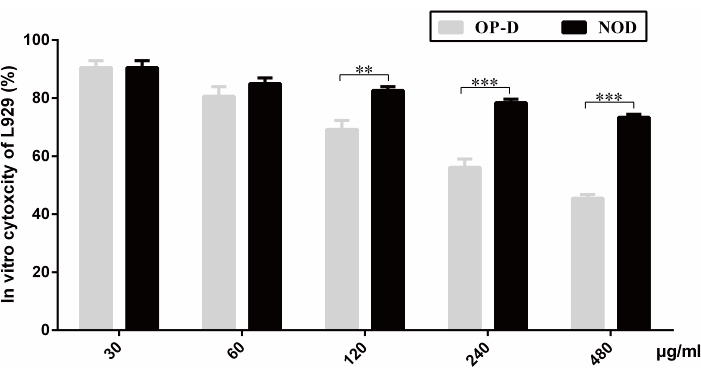
Figure 1: In vitro cytotoxicity test of L929 cells. The cytotoxic effects of different gradient concentrations of OP-D and NOD (30 µg/mL, 60 µg/mL, 120 µg/mL, 240 µg/mL, and 480 µg/mL) when incubated with L929 cells are shown. A CCK-8 kit was used for detection. Statistical analysis was performed using a statistical analysis software. Differences between the two groups were analyzed using an unpaired, two-tailed Student's t-test. All values are expressed as mean ± SD (n = 3), and significant differences are expressed as follows: *p < 0.05, **p < 0.01, and ***p < 0.001. This figure has been modified from Tong et al.14. Please click here to view a larger version of this figure.
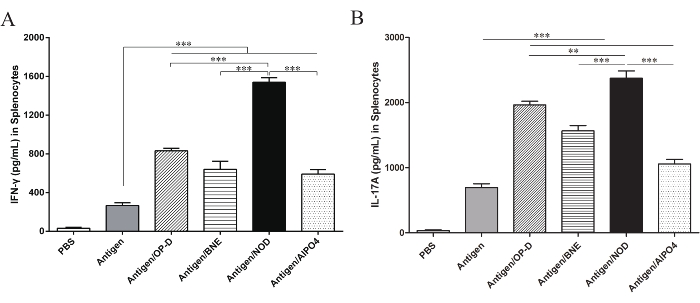
Figure 2: Levels of IFN-γ and IL-17A in splenocytes. Splenocytes from vaccinated mice were stimulated with the antigen for 3 days, and the levels of the cytokines (A) IFN-γ and (B) IL-17A in the supernatant were measured by ELISA. The results showed that IFN-γ and IL-17A production were significantly increased in the Ag/NOD group compared with the Ag/OP-D, Ag/BNE, and Ag/Al groups (p < 0.01). Statistical analysis was performed using a statistical analysis software. Differences among the multiple groups were analyzed using one-way ANOVA followed by Tukey's multiple comparison test. All values are expressed as mean ± SD (n = 8), and significant differences are expressed as follows: *p < 0.05, **p < 0.01, and ***p < 0.001. This figure has been modified from Tong et al.14. Please click here to view a larger version of this figure.
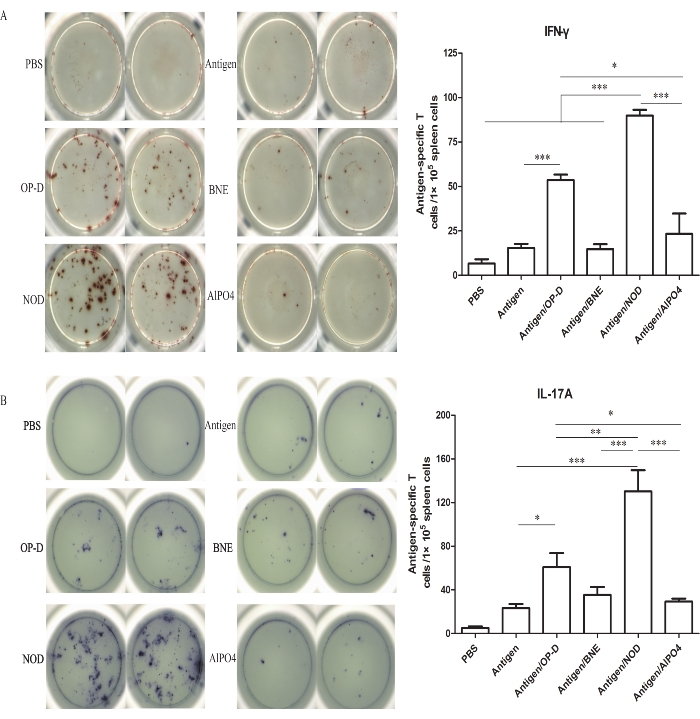
Figure 3: Number of IFN-γ- and IL-17A-secreting cells in the splenocyte population. (A) ELISpot analysis of IFN-γ spot-forming antigen-specific T cells among the splenocytes. (B) ELISpot analysis of IL-17A spot-forming antigen-specific T cells among the splenocytes. Similar to the cytokine results, the Ag/OP-D and Ag/NOD groups showed significantly increased ratios and numbers of IFN-γ- and IL-17A-forming cells in the splenic lymphoid T cell population. The Ag/NOD group induced stronger Th1 (p < 0.001) and Th17 (p < 0.01) immune responses than the other groups. Statistical analysis was performed using a statistical analysis software. Differences among the multiple groups were analyzed using one-way ANOVA followed by Tukey's multiple comparison test. All values are expressed as mean ± SD (n = 8), and significant differences are expressed as follows: *p < 0.05, **p < 0.01, and ***p < 0.001. This figure has been modified from Tong et al.14. Please click here to view a larger version of this figure.
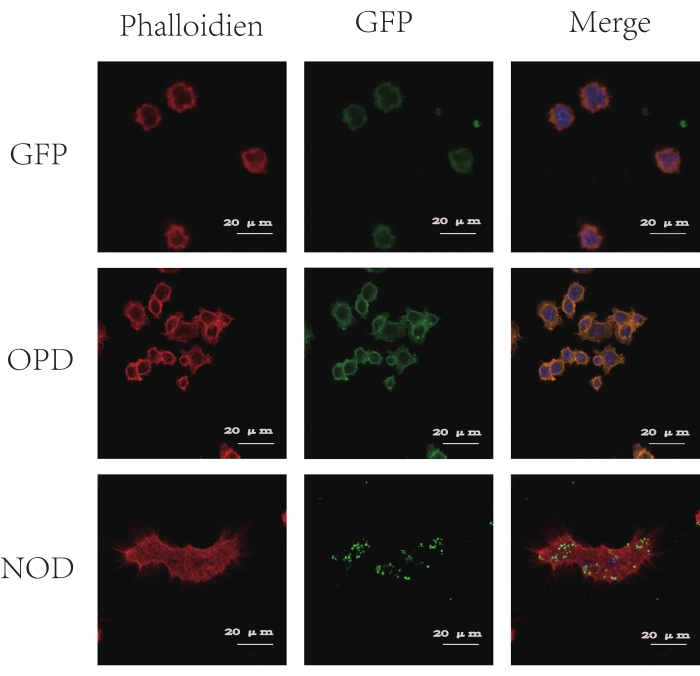
Figure 4: CLSM images of antigen uptake by BMDCs. Phalloidin stains the cytoskeleton in red, DAPI stains the nucleus in blue, and GFP presents green fluorescence. As shown in the figure, green fluorescence is observed in CLSM after 30 min of coincubation of the GFP + OP-D and GFP + NOD groups with BMDCs, but the GFP + NOD particles are surrounded by phagosome-like vesicular structures, while the GFP + OP-D particles are not. This figure has been modified from Tong et al.14. Please click here to view a larger version of this figure.
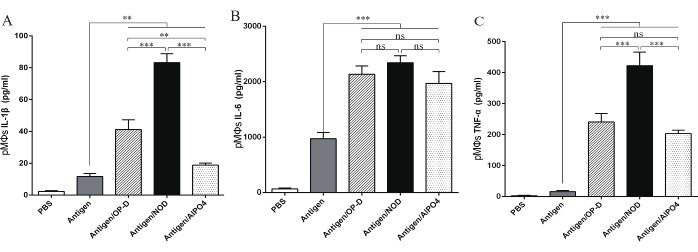
Figure 5: Impact of adjuvant-formulated antigen on the activation of PMs. ELISA for the detection of the concentrations of the cytokines IL-1β, IL-6, and TNF-α in the supernatant of PMs coincubated with PBS, Ag, Ag/OP-D, Ag/NOD, and Ag/Al. Compared with antigen stimulation, Ag/NOD, Ag/OP-D, and Ag/Al stimulation all significantly increased IL-1β, IL-6, and TNF-α secretion from the PMs (p < 0.05). The activation of macrophages was significantly improved in the NOD group compared with the OP-D and AlPO4 groups (p < 0.05). Statistical analysis was performed using a statistical analysis software. Differences among the multiple groups were analyzed using one-way ANOVA followed by Tukey's multiple comparison test. All values are expressed as mean ± SD (n = 8), and significant differences are expressed as follows: *p < 0.05, **p < 0.01, and ***p < 0.001. This figure has been modified from Tong et al.14. Please click here to view a larger version of this figure.
| Reagent | Density | Mass(per 10g) |
| Cremophor EL-35 | bulk | 1.92 |
| glycerol | bulk | 0.48 |
| GTCC | bulk | 0.6 |
| Ultrapure water | bulk | residual amount |
Table 1: The formula of BNE.
| Reagent | Density | Volume |
| β- Mercaptoethanol | 50µM | 0.366µL |
| Fetal Bovine Serum | 1X | 10mL |
| Glutamax | 2mM | 1mL |
| Glutamax RPMI 1640 | 1X | 85mL |
| HEPES | 10mM | 1mL |
| Non-essential amino acids(100x) | 1X | 1mL |
| Penicillin-Streptomycin Solution | 100U/mL | 1mL |
| Sodium pyruvate(100 mM) | 1mM | 1mL |
Table 2: Preparation information for the RF-10 complete medium.
