Tomato (S. lycopersicum L.) is one of the most important vegetable crops around the world, with an output of 186.8 million tons of fleshy fruits from 5.1 million hectares in 20201. It belongs to the large Solanaceae family with about 2,716 species2, including many commercially important crops such as eggplant, peppers, potato, and tobacco. The cultivated tomato is a diploid species (2n = 2x = 24) with a genome size of approximately 900 Mb3. For a long time, great effort has been made toward tomato domestication and breeding by selecting desirable traits from wild Solanum spp. There are over 5,000 tomato accessions listed in the Tomato Genetics Resource Center and more than 80,000 germplasm of tomatoes are stored worldwide4. The tomato plant is perennial in the greenhouse and propagates by seeds. A mature tomato seed consists of three major compartments: a full-grown embryo, residual cellular-type endosperm, and a hard seed coat5,6 (Figure 1A). After double fertilization, the development of cellular-type endosperm precedes the development of zygotes. At ~5-6 days after flowering (DAF), two-celled proembryo is first observed when the endosperm consists of six to eight nuclei7. In Solanum pimpinellifolium, the embryo approaches its final size after 20 DAF, and seeds are viable for germination after 32 DAF8. As the embryo develops, the endosperm is gradually absorbed and only a small amount of endosperm remains in the seed. The residual endosperm consists of micropylar endosperm surrounding the radicle tip, and lateral endosperm in the rest of the seed9,10. The outer seed coat is developed from thickened and lignified outer epidermis of the integument, and with the dead layers of integument remnants, they form a hard shell to protect the embryo and endosperm5.
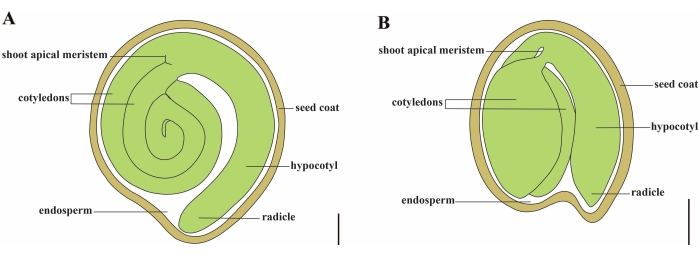
Figure 1: Schematic representation of a mature seed in Solanum lycopersicum and Arabidopsis thaliana. (A) Longitudinal anatomy of a mature tomato seed. (B) Longitudinal anatomy of a mature Arabidopsis seed. A mature tomato seed is approximately 70 times larger in size than an Arabidopsis seed. Scale bars = (A) 400 µm, (B) 100 µm. Please click here to view a larger version of this figure.
Production of high-quality tomato seeds depends on the coordination between the embryo, the endosperm, and the maternal seed components11. Dissecting key genes and networks in seed development requires a deep and full-track phenotypic recording of mutant seeds. Conventional embedding-sectioning techniques, such as the semi-thin section and paraffin section, are widely applied to tomato seeds to observe the local and finer structures of the embryo12,13,14,15. However, analyzing the seed development from thin sections is usually laborious and lacks z-axis spatial resolution. In comparison, tissue clearing is a fast and efficient method to pinpoint the developmental stage of embryo defects that are most likely to occur16. The clearing method reduces the opaqueness of internal tissue by homogenizing the refractive index with one or more biochemical agents16. Whole tissue clearing allows observation of a plant tissue structure without destroying its integrity, and the combination of clearing technology and three-dimensional imaging has become an ideal solution to obtain information on the morphology and developmental state of a plant organ17,18. Over the years, seed clearing techniques have been used in various plant species, including Arabidopsis thaliana, Hordeum vulgare, and Beta vulgaris19,20,21,22,23. Among these, the whole-mount ovule clearing technology has been an efficient approach to studying seed development of Arabidopsis, due to its small size, 4-5 layers of the seed coat cell, and the nuclear-type endosperm24,25. With the continuous updating of different clearing mixtures, such as the emergence of Hoyer's solution26, internal structures of the barley ovule were imaged with a high degree of clarity although its endosperm makes up the bulk of the seeds. Embryogenesis of sugar beet can be observed by clearing combined with vacuum treatment and softening with hydrochloric acid19. Nonetheless, unlike the species mentioned above, embryological observations by clearing protocols in tomato seeds have not been reported. This prevents detailed investigation into the embryonic and seed development of tomatoes.
Chloral hydrate is commonly used as a clearing solution that allows the immersed tissues and cells to be displayed on different optical planes, and substantially preserves the cells or tissue components27,28,29. Chloral hydrate-based clearing protocol has been successfully used for the whole-mount clearing of seeds to observe the embryo and endosperm of Arabidopsis21,28. However, this clearing solution is not efficient in clearing tomato seeds, which are more impermeable than Arabidopsis seeds. The physical barriers include: (1) the tomato integument has nearly 20 cell layers at 3 to 15 DAF30,31, (2) the tomato endosperm is cellular-type, not nuclear-type32, and (3) tomato seeds are about 70 times larger in size33,34 and (4) produce large amounts of seed coat mucilage, which blocks the penetration of clearing reagents and affects the visualization of embryo cells.
Therefore, this report presents an optimized chloral hydrate-based clearing method for whole-mount clearing of tomato seeds at different stages, which allows deep imaging of the embryo development process (Figure 2).
