G-Protein-gekoppelte Rezeptoren (GPCRs), die von der Hefe bis zum Menschen vorhanden sind, stellen die größte Superfamilie von Rezeptoren in vielen Organismen dar1. Sie spielen eine entscheidende Rolle bei der Regulierung fast aller biologischen Prozesse bei Tieren. Es gibt 50-200 GPCRs im Genom von Arthropoden, was bedeutet, dass sie die größte Membranrezeptor-Superfamilie2 darstellen. Sie werden in sechs Hauptklassen, A-F, eingeteilt, basierend auf ihrer Sequenzähnlichkeit und ihren Funktionen3. GPCRs transduzieren verschiedene extrazelluläre Signale, wie z.B. die von Hormonen, Neuropeptiden, biogenen Aminen, Glutamat, Protonen, Lipoglykoproteinen und Photonen4. GPCRs koppeln an heterotrimere G-Proteine (Gα, Gβ und Gγ), um nachgeschaltete Signale zu übertragen. GPCRs, die an Gαs oder Gαi/o-Proteine gekoppelt sind, erhöhen bzw. senken die intrazellulären 3′, 5′-zyklischen Adenosinmonophosphat (cAMP)-Spiegel durch Aktivierung oder Hemmung der Adenylylcyclase. GPCRs, die an Gαq/11 gekoppelt sind, induzieren die Freisetzung von Kalzium aus den Kalziumspeichern des endoplasmatischen Retikulums, indem sie den Phospholipase C (PLC)-Inositol-1,4,5-triphosphat (IP3) -Weg aktivieren. GPCRs, die an Gα12/13 gekoppelt sind, aktivieren die RhoGTPase-Nukleotidaustauschfaktoren 5,6. GPCRs sind das Ziel von mehr als 50% der Humanarzneimittel und eines Akarizids, Amitraz4. Da GPCRs so unterschiedliche Signale übertragen, sind sie vielversprechende Ziele für die Entwicklung neuartiger Pestizide, die wirbellose physiologische Funktionen stören.
Das Ziel von HTS ist es, Hit-Moleküle zu identifizieren, die Rezeptorfunktionen modulieren können. HTS umfasst Assay-Entwicklung, Miniaturisierung und Automatisierung7. Arthropoden-Neuropeptid-GPCRs sind an den meisten physiologischen Funktionen wie Entwicklung, Häutung und Ekdysis, Ausscheidung, Energiemobilisierung und Fortpflanzungbeteiligt 4. Die meisten Neuropeptid-GPCRs von Arthropoden und Metazoen signalisieren durch die Calcium-Signalkaskade 2,6,8,9,10, wie z.B. in den myoinhibitorischen Peptid- und SIFamid-Rezeptoren der schwarzbeinigen Zecke Ixodes scapularis; Ihre Liganden sind in den Motilitätstests des Hinterdarms antagonistisch, wobei SIF eine Kontraktion auslöst und MIP sie hemmt11,12. Ein NPY-ähnlicher Rezeptor der Gelbfiebermücke, Aedes aegypti, reguliert die weibliche Wirtssuche13. Im Vergleich zu anderen alternativen Calcium-Mobilisierungsassays, wie dem Aequorin-Calcium-Biolumineszenz-Assay14, ist der Calcium-Fluoreszenz-Assay einfach durchzuführen, erfordert keine Transfektion anderer rekombinanter Calcium-Detektionsproteine und ist kostengünstig. Der Calcium-Fluoreszenz-Assay erzeugt ein verlängertes Signal im Vergleich zu dem schnellen kinetischen Signal, das im Aequorin-Calcium-Biolumineszenz-Assay14,15 erhalten wurde.
Im vorliegenden Beispiel wurde der Kinin-Rezeptor aus der Rinderpestzecke Rhipicephalus microplus rekombinant in der CHO-K1-Zelllinie exprimiert und für den Calciumfluoreszenz-Assay verwendet. Es gibt nur ein Kinin-Rezeptor-Gen in R. microplus; Der Rezeptor signalisiert über einen Gq-Protein-abhängigen Signalweg und löst den Ausfluss von Ca2+ aus Calciumspeichern in den intrazellulären Raum16 aus. Dieser Prozess kann durch ein Fluorophor nachgewiesen und quantifiziert werden, das bei der Bindung von Calciumionen ein Fluoreszenzsignal auslöst (Abbildung 1).
Der Kinin-Rezeptor ist ein wirbellosen-spezifischer GPCR, der zu den Klasse-A-Rhodopsin-ähnlichen Rezeptoren gehört. Kinin ist ein uraltes Signal-Neuropeptid, das in Mollusca, Crustacea, Insecta und Acari 4,17,18 vorkommt. Coleopteren (Käfer) fehlt das Kinin-Signalsystem; In der Mücke Aedes aegypti gibt es nur einen Kinin-Rezeptor, der drei Aedeskinine bindet, während Drosophila melanogaster einen Kinin-Rezeptor mit Drosokinin als einzigartigem Ligandenhat 19,20,21. Bei Wirbeltieren gibt es keine homologen Kinine oder Kininrezeptoren. Obwohl die genaue Funktion von Kinin bei Zecken unbekannt ist, zeigen die Kininrezeptor-RNAi-stillgelegten Weibchen von R. microplus eine signifikant reduzierte Fortpflanzungsfähigkeit22. Kinine sind pleotrope Peptide in Insekten. Bei Drosophila melanogaster sind sie sowohl am zentralen als auch am peripheren Nervensystembeteiligt 23, an der Präekdysis24, an der Nahrungsaufnahme25, am Stoffwechsel 26 und an den Schlafaktivitätsmustern 26,27 sowie an der Fortbewegung der Larven 28. Kinine regulieren die Kontraktion des Hinterdarms, die Diurese und die Fütterung der Mücke A. aegypti 29,30,31. Die Kininpeptide weisen ein konserviertes C-terminales Pentapeptid Phe-X1-X2-Trp-Gly-NH2 auf, das die minimal erforderliche Sequenz für die biologische Aktivität32 darstellt. Die Arthropodenspezifität, die geringe Größe des endogenen Liganden, die sie für kleinmolekulare Interferenzen empfänglich macht, und die pleiotropen Funktionen bei Insekten machen den Kininrezeptor zu einem vielversprechenden Ziel für die Schädlingsbekämpfung4.
Der “Dual-Addition”-Assay (Abbildung 2) ermöglicht die Identifizierung von Agonisten oder Antagonisten im selben HTS-Assay15. Es basiert auf einem “Dual-Addition”-Assay, der in der pharmazeutischen Industrie häufig für die Wirkstoffforschung verwendet wird33. Kurz gesagt, die erste Zugabe von Medikamenten in die Zellplatte ermöglicht die Identifizierung potenzieller Agonisten in der chemischen Bibliothek, wenn ein höheres Fluoreszenzsignal im Vergleich zur Anwendung der Lösungsmittelkontrolle detektiert wird. Nach 5 Minuten Inkubation mit diesen kleinen Molekülen wird ein bekannter Agonist (Kinin-Peptid) auf alle Vertiefungen aufgetragen. Diejenigen Vertiefungen, die zufällig einen Antagonisten von der Wirkstoffplatte erhielten, zeigen ein niedrigeres Fluoreszenzsignal bei der Agonistenzugabe im Vergleich zu den Kontrolltöpfen, die das Lösungsmittel bei der ersten Zugabe erhielten. Dieser Assay ermöglicht dann die Identifizierung potenzieller Agonisten und Antagonisten mit den gleichen Zellen. In einem Standard-HTS-Projekt würden diese Hit-Moleküle durch Dosis-Wirkungs-Assays und durch zusätzliche biologische Aktivitätsassays, die hier nicht gezeigt werden, weiter validiert.
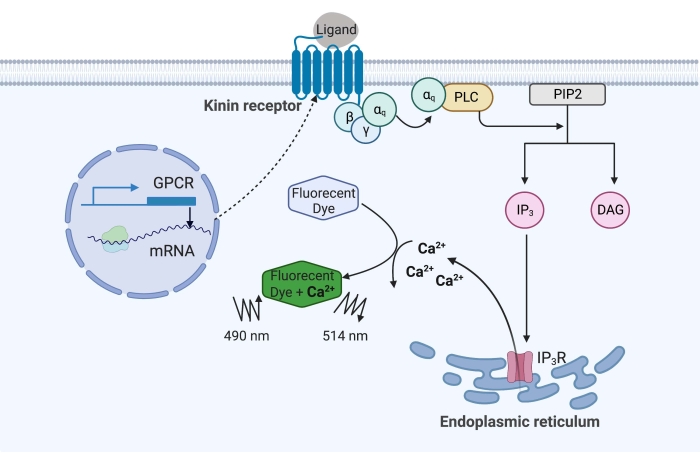
Abbildung 1: Illustration des Calciumfluoreszenz-Assay-Mechanismus. Das Gq-Protein löst den intrazellulären Kalzium-Signalweg aus. Der Kinin-Rezeptor (G-Protein-gekoppelter Rezeptor) wurde in CHO-K1-Zellen rekombinant exprimiert. Wenn der Agonistenligand an den Rezeptor bindet, aktiviert das mit dem Kininrezeptor assoziierte Gq-Protein PLC, das die Umwandlung eines PIP-2-Moleküls in IP3 und DAG katalysiert. IP 3 bindet dann an die IP3R auf der Oberfläche des endoplasmatischen Retikulums, was zur Freisetzung von Ca 2 + in das Zytoplasma führt, wo Ca2 + -Ionen an die Fluorophore binden und ein Fluoreszenzsignal auslösen. Das Fluoreszenzsignal kann durch Anregung bei 490 nm erhalten und bei 514 nm detektiert werden. Abkürzungen: GPCR = G-Protein-gekoppelter Rezeptor; PLC = phospholipase C; PIP2 = Phosphatidylinositol-4,5-bisphosphat; IP3 = Inositoltrisphosphat; DAG = Diacylglycerin; IP 3 R =IP-3-Empfänger. Erstellt mit BioRender.com. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
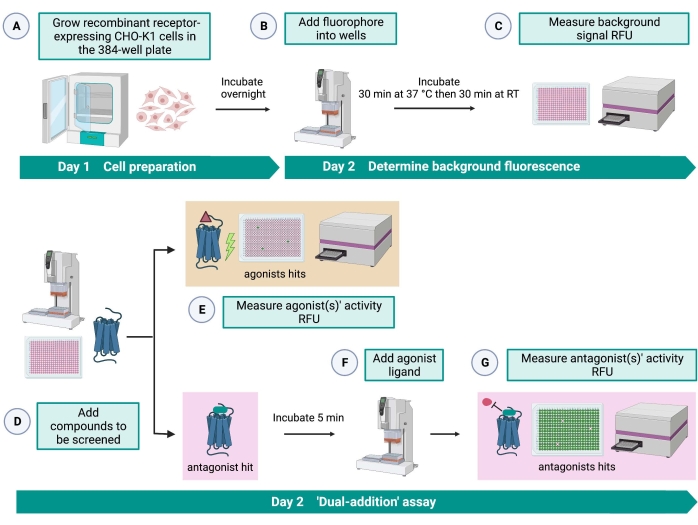
Abbildung 2: Der Arbeitsablauf für das Hochdurchsatz-Screening von kleinen Molekülen auf einem G-Protein-gekoppelten Rezeptor, der in CHO-K1-Zellen exprimiert wird. (A) Rekombinante CHO-K1-Zellen, die den Kinin-Rezeptor stabil exprimieren, wurden unter Verwendung eines Liquid-Handling-Systems (25 μL/Well) in die 384-Well-Platte (10.000 Zellen/Well) gegeben und in einem befeuchteten CO2 -Inkubator für 12-16 h inkubiert. ) Der Assay-Puffer, der den Fluoreszenzfarbstoff (25 μl/Well) enthielt, wurde unter Verwendung eines Liquid-Handling-Systems in die Zellplatte gegeben. Die Platte wurde für 30 min bei 37 °C für 30 min inkubiert und bei RT für weitere 30 min äquilibriert. (C) Das Hintergrundfluoreszenzsignal der Zellen in jeder Vertiefung wurde mit einem Plattenleser gemessen. (D) Arzneimittellösungen aus einer Bibliotheksplatte mit 384 Vertiefungen und ein Leerlösungsmittel (alle mit 0,5 μl/Well) wurden unter Verwendung eines Liquid-Handling-Systems in die zelluläre Assayplatte gegeben. (E) Die zellulären Calciumfluoreszenzreaktionen wurden unmittelbar nach der Zugabe der Arzneimittellösungen mit dem Plattenleser gemessen; Verbindung(en), die überdurchschnittliche Fluoreszenzsignale hervorruft, wurden als Agonisten-Treffer ausgewählt. Antagonisten-Treffer, die die GPCR blockieren (Symbol unten), wurden nach der Zugabe des Peptidagonisten während Schritt G aufgedeckt. (F) In der gleichen Testplatte wurde nach 5 min Inkubation der Zellen mit Screening-Verbindungen ein endogenes Agonistenpeptid Rhimi-K-1 (QFSPWGamid) des Zeckenkininrezeptors zu jeder Vertiefung (1 μM) gegeben. (G) Zelluläre Fluoreszenzreaktionen nach der Agonisten-Peptidaddition wurden vom Plattenleser sofort gemessen. Verbindung(en), die das Fluoreszenzsignal hemmen, wurden als Antagonisten-Treffer ausgewählt. Abkürzungen: GPCR = G-Protein-gekoppelter Rezeptor; RT = Raumtemperatur; RFU = relative Fluoreszenzeinheiten. Erstellt mit BioRender.com. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
